papillae contains bundles of 20–27 µm long extrusomes
vermiform macronucleus
contractile vacuole terminal
Legendrea pespelicani
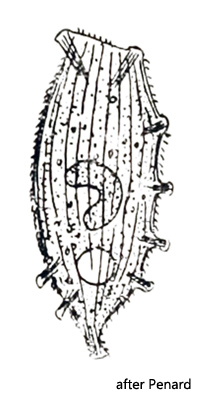
The first description of Legendrea pespelicani by Penard was based on the finding of a single specimen. Although Penard lived to be 90 years old and certainly examined thousands of specimens, this one encounter remained the only one. No other finds have been documented since. Also Kahl took over 1935 only the description of Penard. In October 2007, I found a total of 3 specimens of Legendrea pespelicani in the Simmelried in the uppermost layers of the sludge at a depth of about 50 cm and 7 years later, in July 2014, another 6 specimens.
Legendrea pespelicani was first described by Penard. The genus Legendrea includes ciliates that are closely related to the Spathidia. Like the Spathidia, they have a spatulate oral bulge that is equipped with extrusomes. However, they are distinguished from spathidia by the possession of papillae-shaped bundles of extrusomes distributed over the body along an imaginary linear extension of the oral bulge (meridional arrangement). It is assumed that these bundles of extrusomes are a result of a segmentation and partitioning of the oral bulge. These separate bundles may either located at the end of tentacles (as in Legendrea loyezae, for example) or as papillae-like elevations on the cell body. Later, Jankowski (1967) transferred Legendrea pespelicani to the genus Lacerus. However, it turned out that the genus Lacerus is invalid, so the name Legendrea pespelicani originally given by Penard is the currently valid name.
The specimen I have found in 2007 were distinctly laterally flattened. All three specimens had 12 papillae, arranged on the dorso-ventral line, protruding visibly from the cell body (s. fig. 1 a-c).
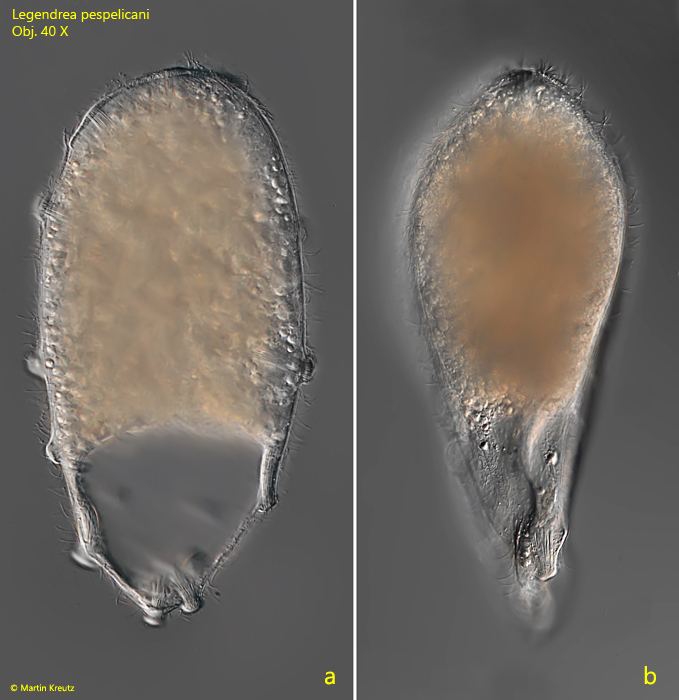
Fig. 1 a-c:Legendrea pespelicani. L = 305 µm. A freely swimming specimen. The 12 papillae ( nos. 1–12) are clearly visible. Obj. 40 X.
It almost seemed as if the cell membrane was stretched between these papillae. This may be a reason for the species name pespelicani (= foot of the pelican) by Penard. Perhaps Penard felt reminded of a webbed skin stretched between the toes of the pelican or of a correspondingly shaped shell with the name “pelican foot” (Aporrhais pespelecani).
The 6 specimens found 7 years later differed markedly from this body shape. They seemed to be well-fed specimens, which were almost rotund in the anterior half of the body and only the posterior end was flattened (s. fig. 2 a-b).
Fig. 2 a-b:Legendrea pespelicani. L = 240 µm. A second freely swimming specimen in lateral (a) and dorsal view (b). Obj. 40 X.
The well-fed “thicker” specimens could be easily confused with Holophrya ovum at small magnifications, which was also found in the specimens because the papillae did not protrude so clearly on the body margin.
I could examine the arrangement of the characteristic papillae of Legendrea pespelicani best on the dorso-ventrally flattened specimens of 2007. The number and position of the papillae on the ventral and dorsal edge of the cell body could be determined very well. In the Simmelried population, there are constantly 12 of these papillae, which, when counted clockwise, starting from the apically located papilla, are arranged as follows (s. fig. 1a):
This finding is not in accordance with the data and drawings of Penard (1922), who gives 10 papillae for this species. The terminal papillae 8 and 9 are characterized by an apparently obliquely angled arrangement, which is clearly visible in lateral view (s. fig. 2a). It is caused by the underlying contractile vacuole. Due to its volume, the overlying plasma layer is very thin and the extrusomes of papillae 8 and 9 cannot arrange themselves perpendicularly to the cell center as a result. As a result, they seem to be arranged somewhat like “crooked teeth”. In a dorsal view, I could see that the arrangement of the papillae follows the course of the meridional dorsal line quite closely. The papillae are about 15–18 µm in diameter and are formed by dense bundles of 20–27 µm long, slightly curved extrusomes that are embedded into the cytoplasm.
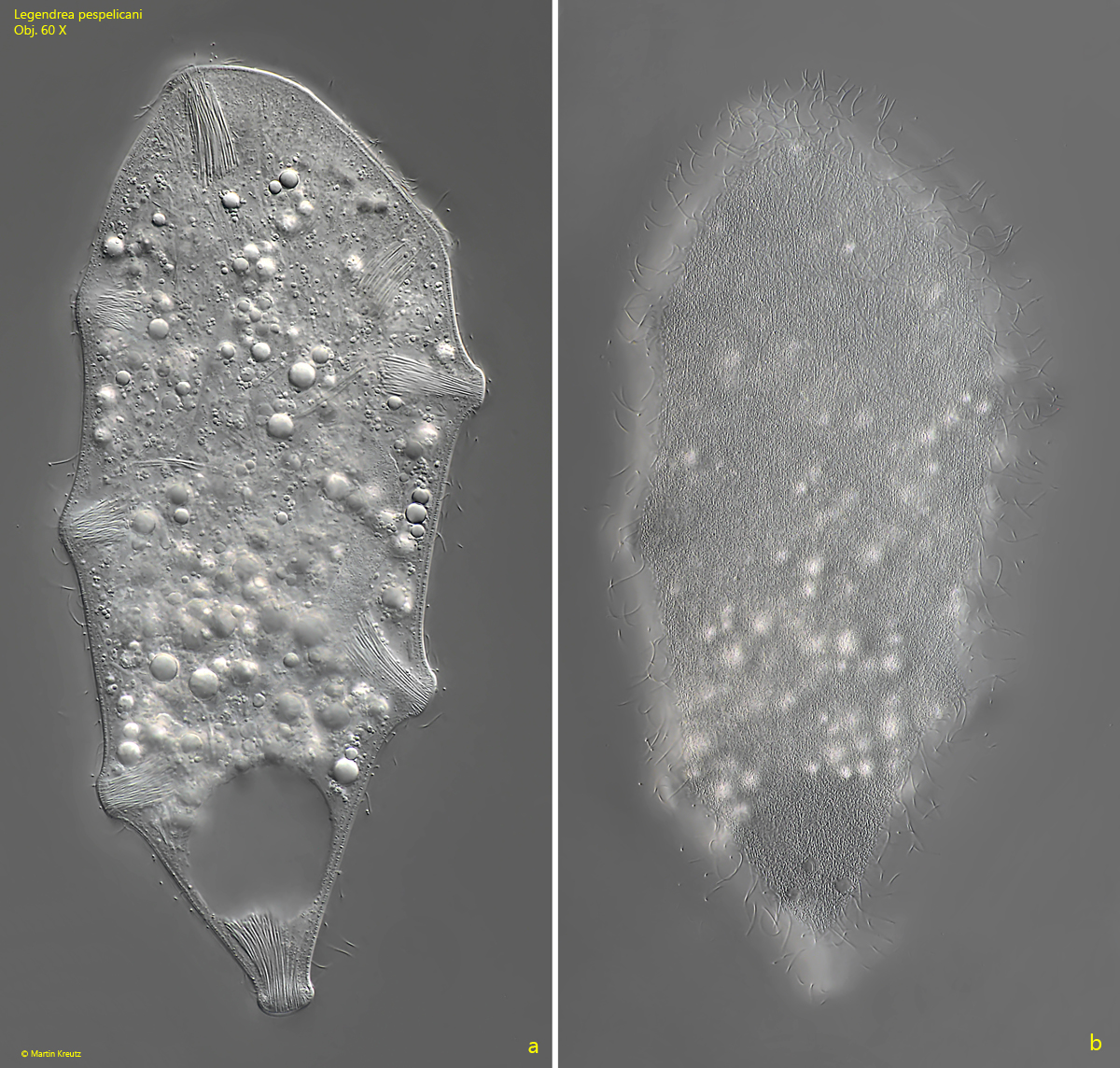
In August 2023 I found another 5 specimens in the Simmelried. All of them had only 8 papillae (s. fig. 3 a-b). Thus their number seems to be quite variable. The range of 8–12 papillae includes also the finding of Penard at that time, who counted 10 papillae.
Fig. 3 a-b:Legendrea pespelicani. L = 245 µm. Two focal planes of slightly squashed specimen with only 8 papillae. Obj. 60 X.
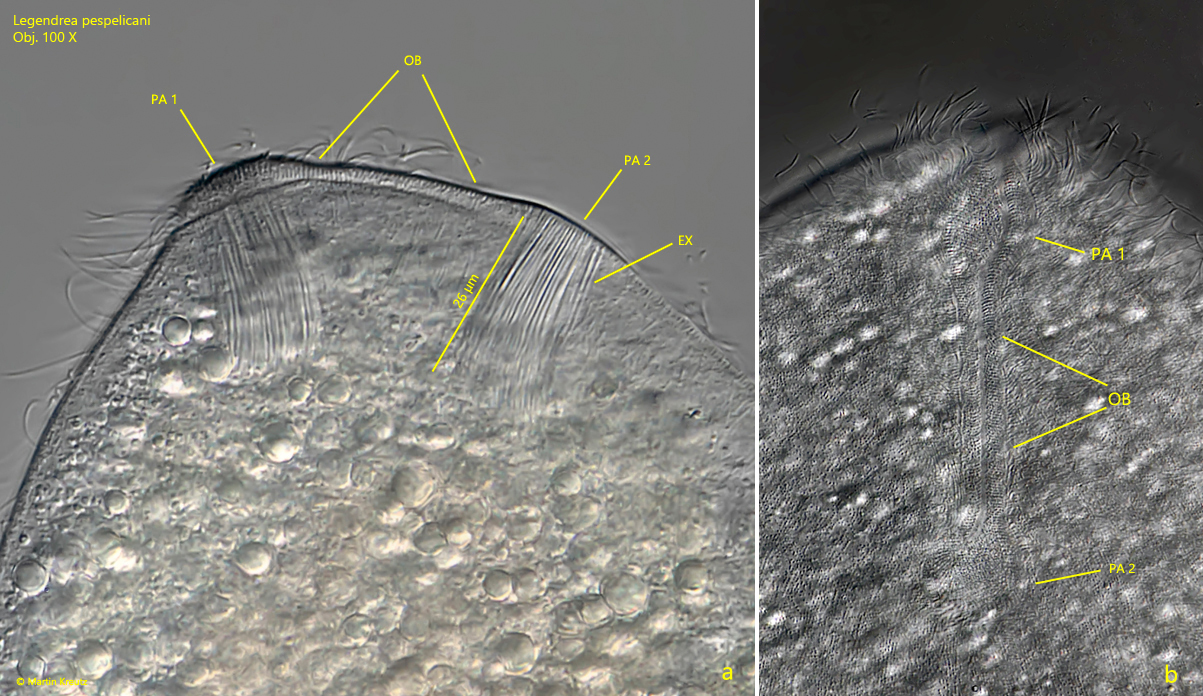
The apical oral bulge is not easy to observe, but according to my observations it is bounded by papillae 1 and 2, which are also part of the oral bulge (s. fig. 4 a-b).
Fig. 4 a-b:Legendrea pespelicani. Lateral (a) and ventral view (b) on the oral bulge (OB), located between papillae 1 and 2 (PA 1, PA 2). Obj. 100 X.
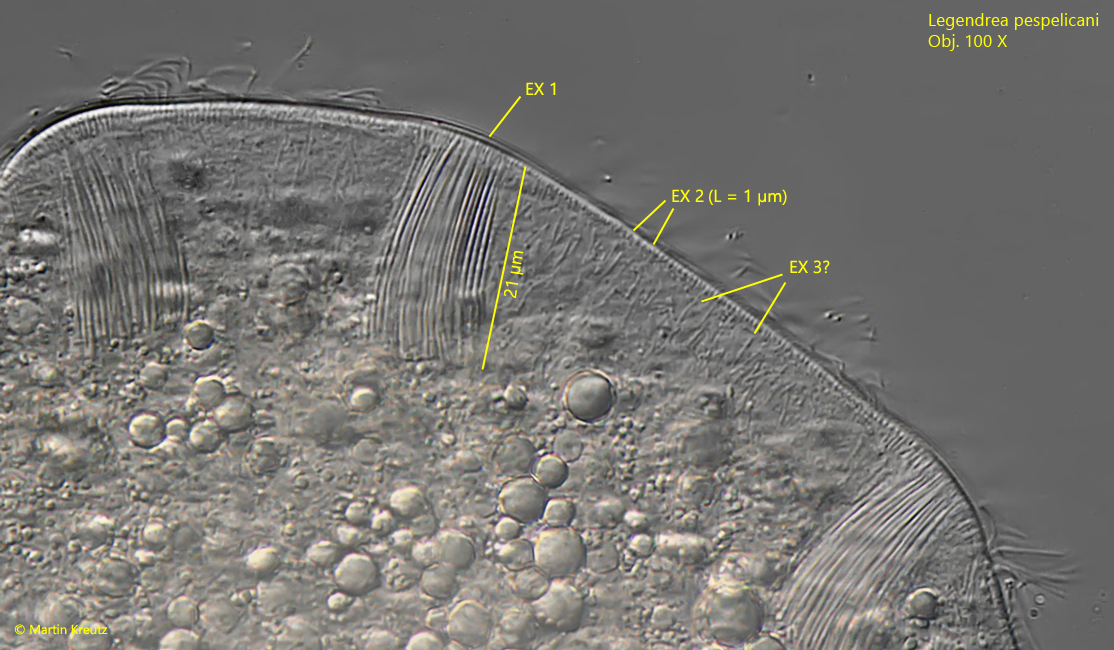
The extrusomes in the papillae are 20–27 µm long, slightly curved and tapered towards one end (s. figs. 5, 6 and 7). Beside these prominent extrusomes there is a fringe of short rods with a length of about 1 µm. A third type of extrusomes could not be clearly identified, as it could also be symbiotic bacteria in the cytoplasm (s. fig. 5).
Fig. 5:Legendrea pespelicani. The prominent extrusomes in the papillae (EX 1) are 20 – 27 µm long. A second type of extrusomes (EX 2) is located beneath the pellicle. These extrusomes are about 1 µm long rods. The presence of a third type of extrusomes (EX 3) is not confirmed because it could be also symbiontic bacteria. Obj. 100 X.
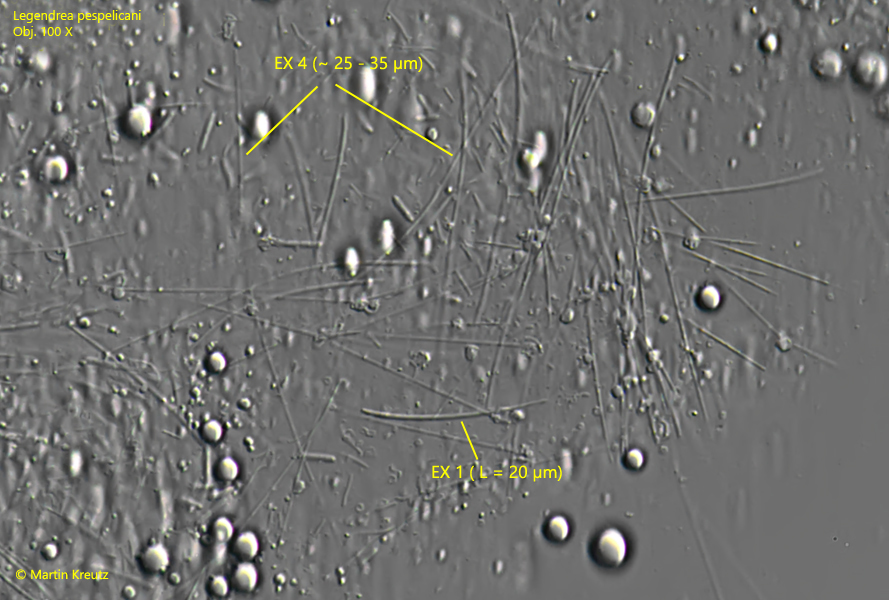
In a strongly squashed specimen a further type of extrusomes could be identifiend (EX 4, s. fig. 6). These extrusomes are 25–35 µm long thin rods. In total, there are at least 3 types of extrusomes present in Legendrea pespelicani.
Fig. 6:Legendrea pespelicani. In a strongly squashed specimen thin rods with a length of 25–35 could be identified as an further type of extrusomes (EX 4). The prominent extrusomes in the papillae (EX 1) are curved rods with one tapered end. Obj. 100 X.
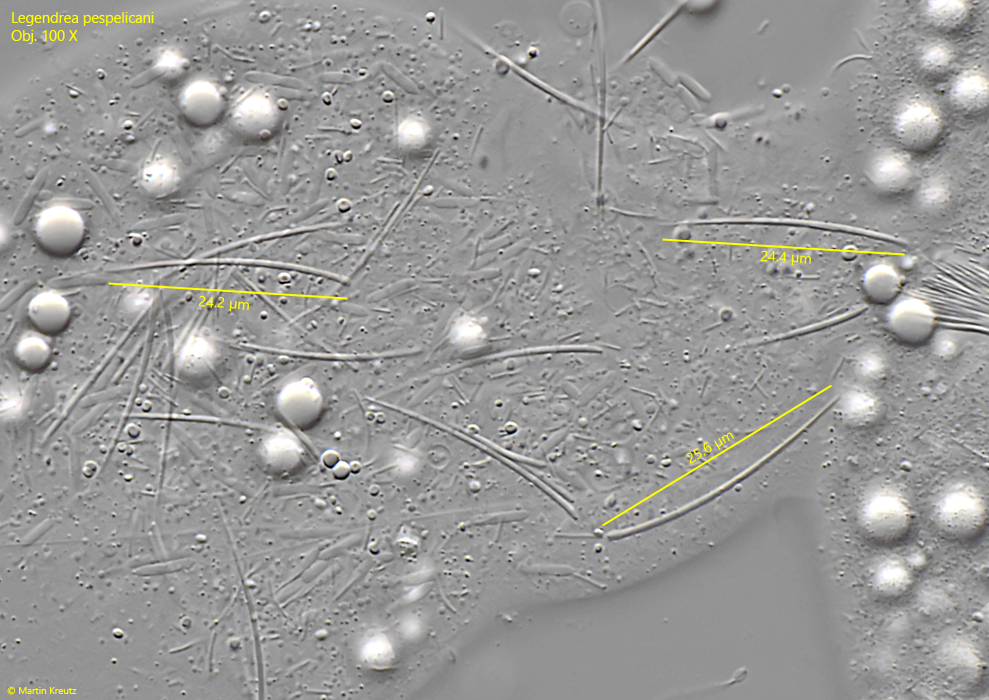
Fig. 7:Legendrea pespelicani. The extrusomes EX 1 in a second specimen with a length of 24–25 µm. Obj. 100 X.
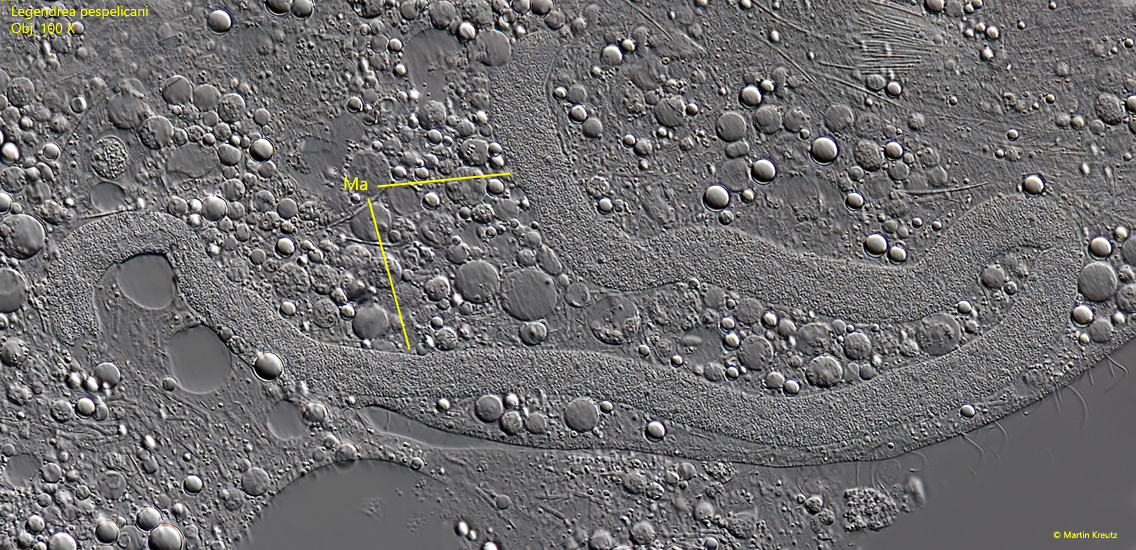
The body length of all 15 specimens found between 2007 – 2023 was between 240 and 310 µm and thus about 30 % larger than Penard’s specimen, for which he gives a length of 210 µm. In all specimens the body was filled with spheres of oily reserve substances. The contracile vacuole is large and located terminally. Only in squeezed specimens the 200–250 µm long and 7–10 µm thick, vermiform macronucleus could be seen well (s. fig. 8). I could not identify the micronucleus, due to the reserve substances.
Fig. 8:Legendrea pespelicani. The vermiform macronucleus (Ma) of this specimen is 350 µm long. Obj. 100 X.
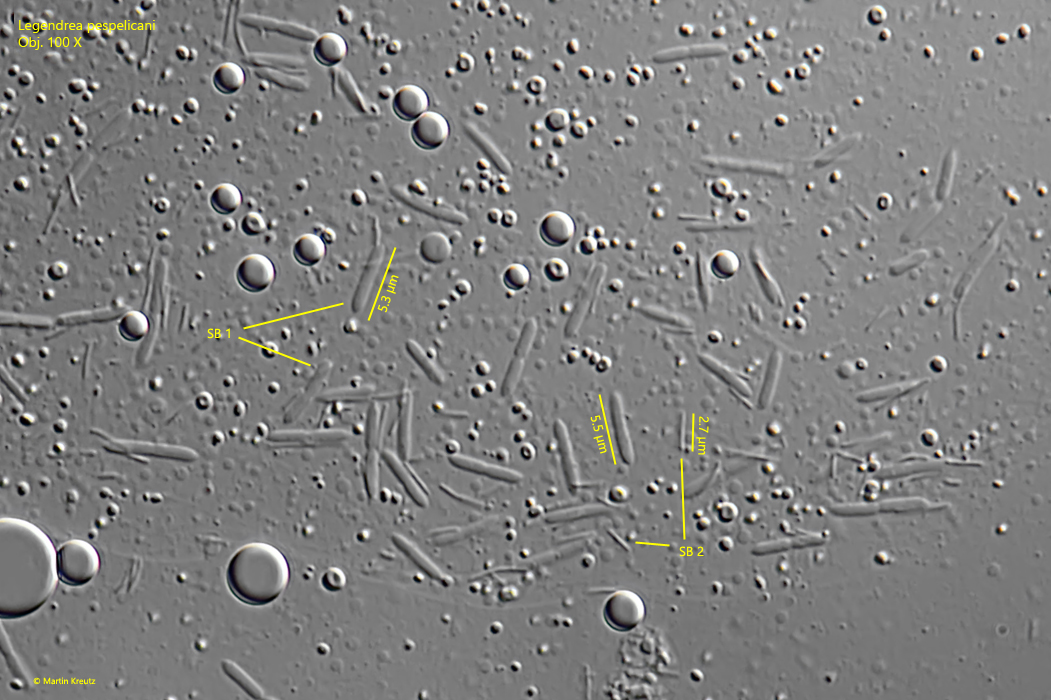
Two different symbiotic bacteria were found in the cytoplasm of Legendrea pespelicani. The smaller bacteria were rods with a length of 2.5–3 µm, while the larger bacteria were oblong with slightly pointed ends and a length of 5–6 µm (s. fig. 9).
Fig. 9:Legendrea pespelicani. There are two types of symbiotic bacteria (SB 1, SB 2). The larger ones (SB 1) are oblong rods with slightly tapered ends and a length of 5–6 µm, while the small bacteria (SB 2) are thin rods with a length of 2.5–3 µm. Obj. 100 X.
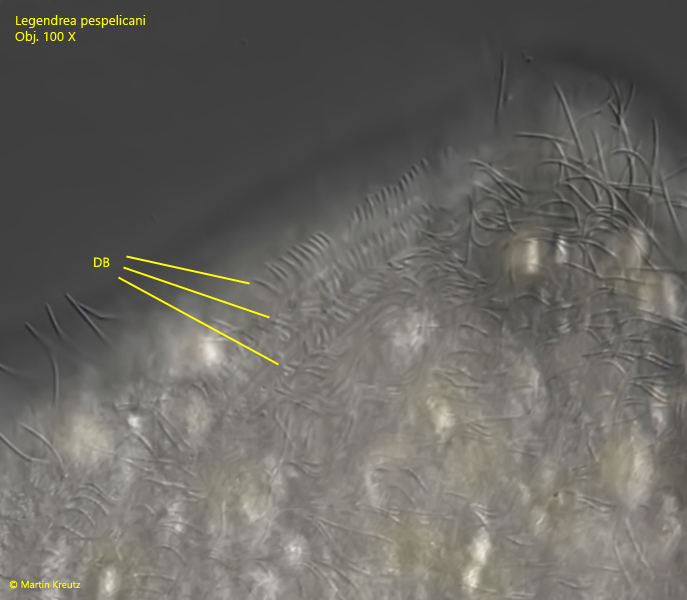
A three-rowed dorsal brush with bristles about 3.5 µm long is located dorsally to the oral bulge (s. fig. 10).
Fig. 10:Legendrea pespelicani. The dorsal brush consists of three rows of short cilia with a length of approx. 3.5 µm. Obj. 100 X.
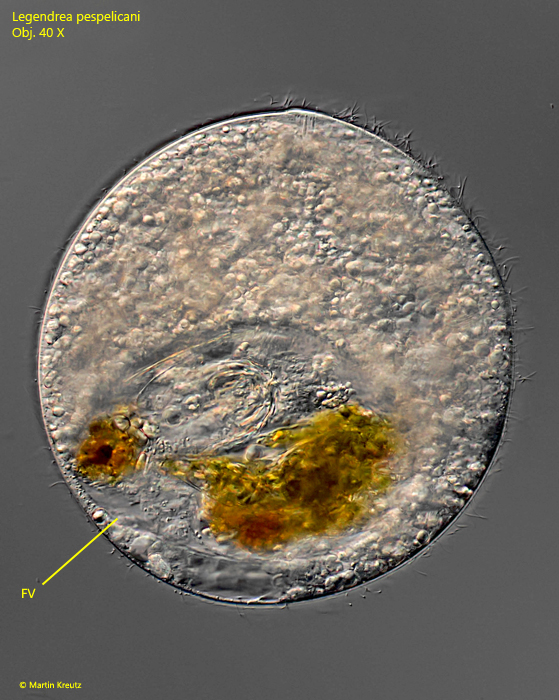
Unfortunately, I was never able to observe feeding in all 9 specimens of Legendrea pespelicani to learn about the role of the papillae. Only in one specimen I could find a filled food vacuole (s. fig. 11).
Fig. 11:Legendrea pespelicani. The only specimen found with a filled food vacuole (FV). Obj. 40 X.
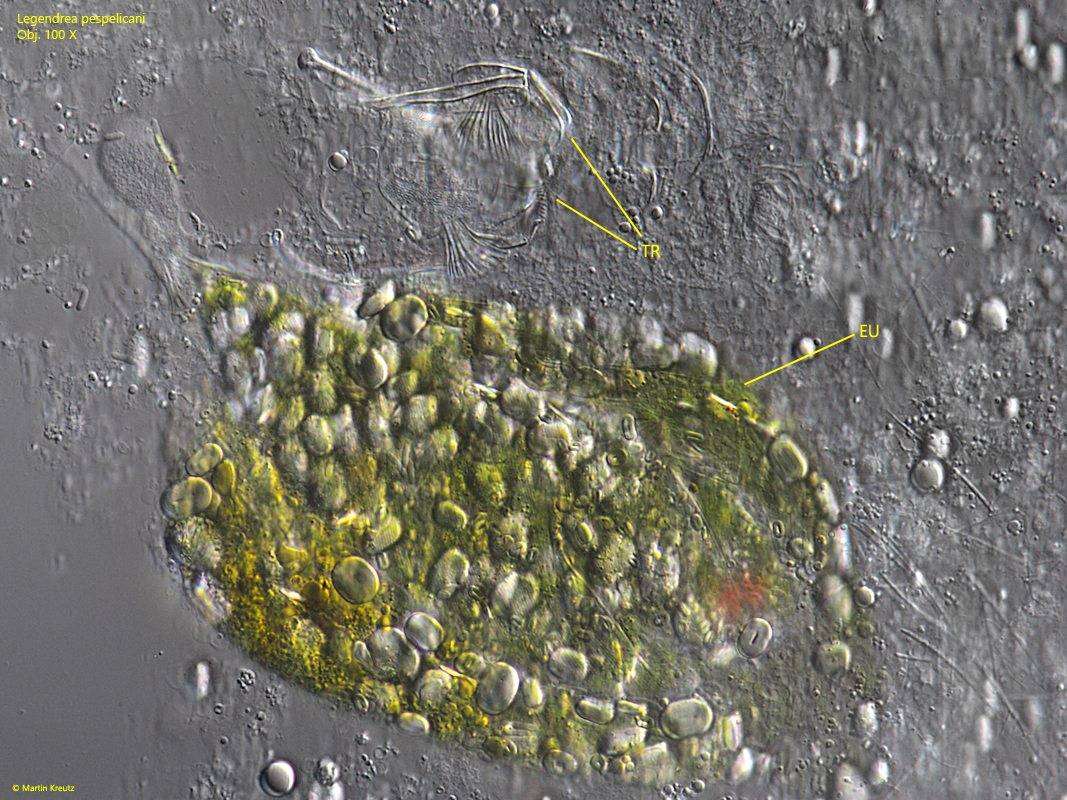
To examine the contents of the food vacuole more closely, I squeezed the specimen. I found the remains of at least three prey organisms (s. fig. 12). One was a rotifer, which could be assigned to the genus Cephalodella (probably C. giganthea or C. tenuiseta) on the basis of the undigested trophi. In addition, the remains of at least two specimens of Euglena sanguinea could be identified, based on the shape of the paramylon granules and the blunt terminal spine. Both possible species of Cephalodella and Euglena sanguinea were frequently found in the sample. Thus, it is obvious that Legendrea pespelicani is not a food specialist and also devours large prey.
Fig. 12:Legendrea pespelicani. The content of the food vacuole were an undigested trophi of a rotifer (likely Cephalodella gigantea or Cephalodella tenuiseta) and remains of two specimens of Euglena sanguinea. Obj. 100 X.