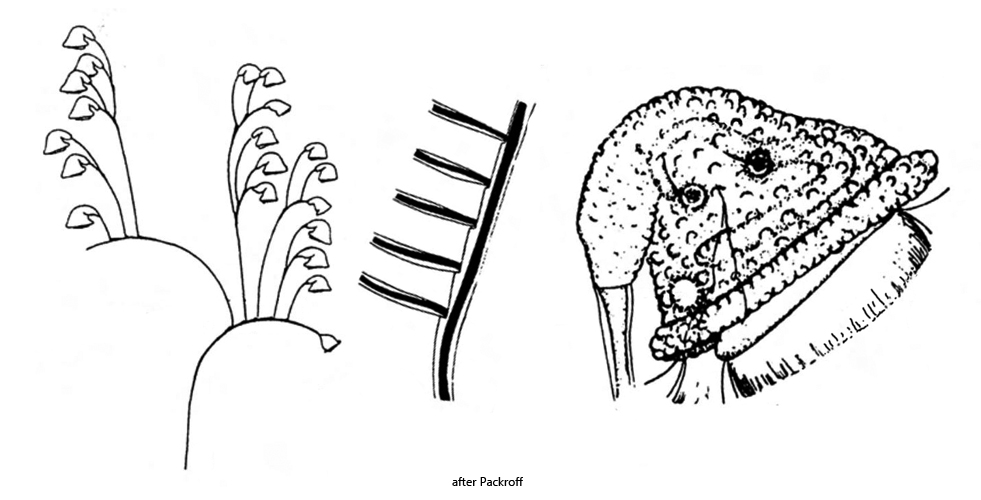
extended zooids campanulate, nodding at distal end of stalks
contracted zooids globular
length of extended zooids 40–70 µm
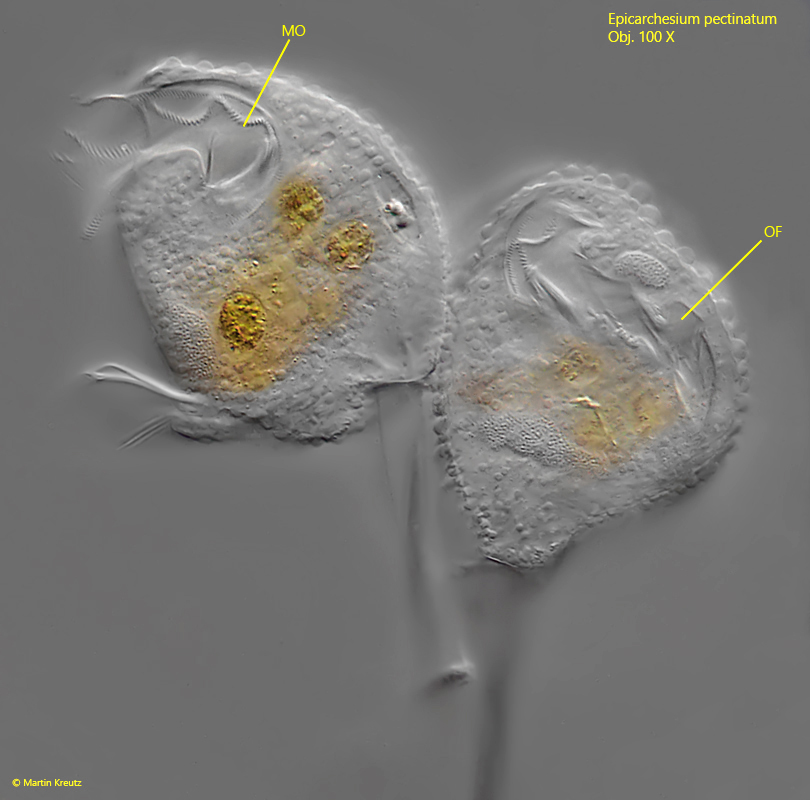
peristomal collar projecting body slightly
macronucleus J–shaped
two contractile vacuoles
pellicle covered with tubercles
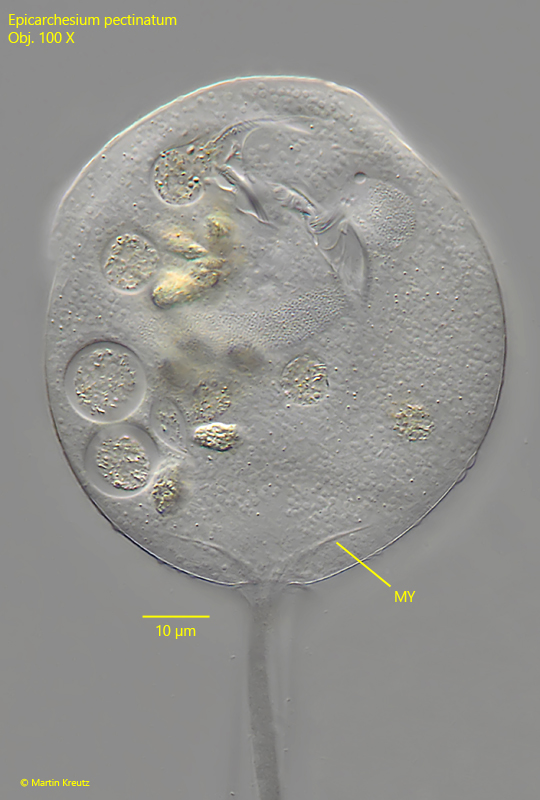
stalk with distinct myoneme, contracts spirally
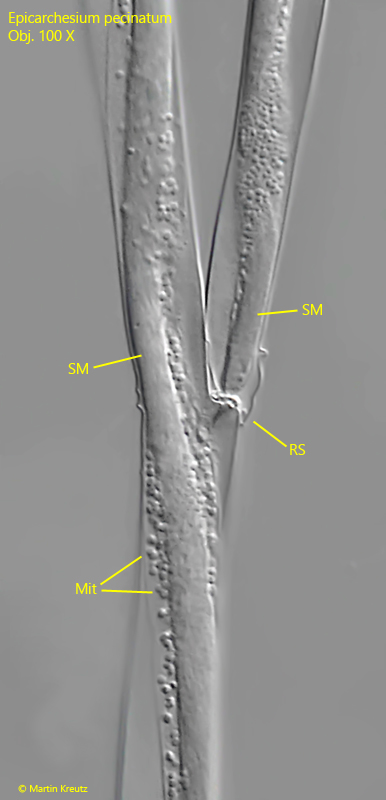
stalk branched, myonems interrupted at ramification sites
serrated structure of stalk due to detached daughter cells
Epicarchesium pectinatum
Epicarchesium pectinatum was first described by Zacharias (1897) as Zoothamnium pectinatum. After a re-description by Kahl (1935), the species was renamed Carchesium pecitinatum. However, important features of the species are a tuberculate pellicle and the myonema in the branched stalks are interrupted at the ramification sites. On the basis of these features, Foissner, Berger and Schaumburg (1999) finally placed the species in the genus Epicarchesium.
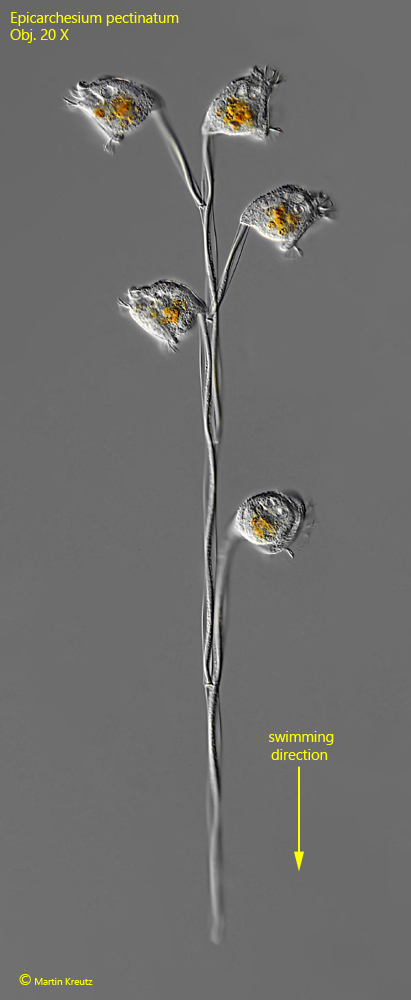
I have only found Epicarchesium pectinatum once so far, in May 2020. The site was the eutrophic pond of the waste disposal company Constance. The colonies in my samples consisted of up to 20 zooids. I did not find any larger colonies. The first characteristic for identification is the swimming style, because the colonies swim with the stem end first (s. fig. 1).
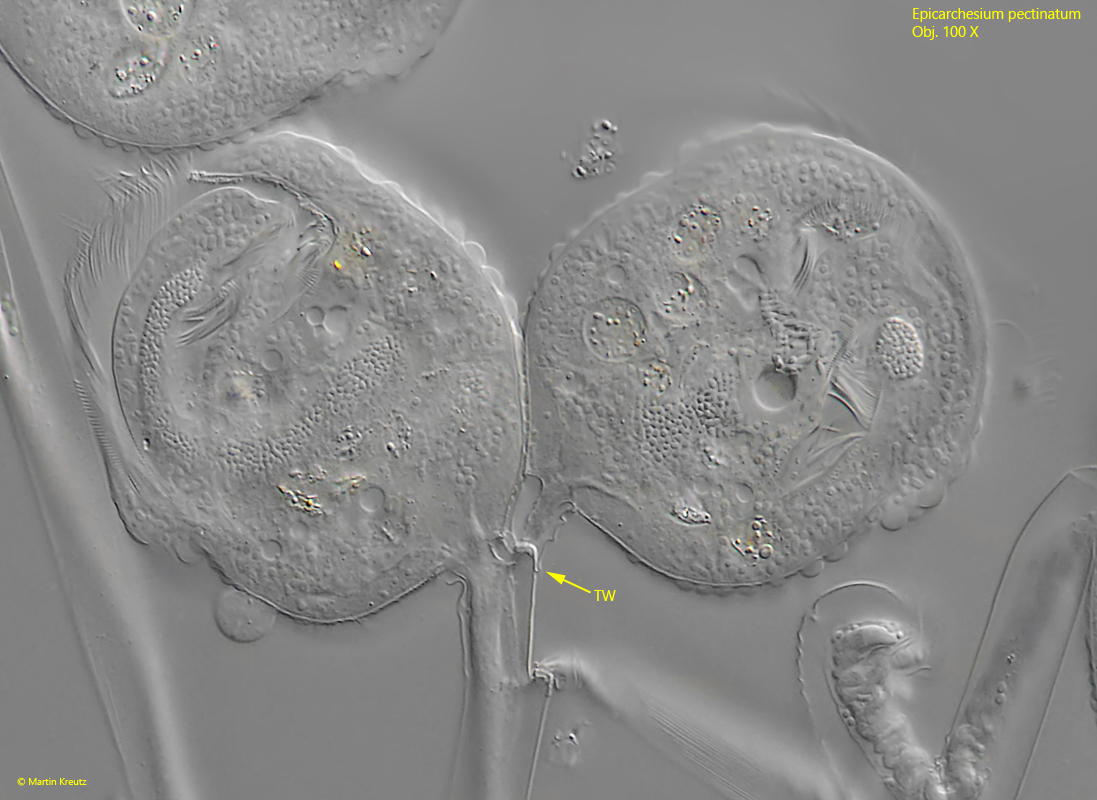
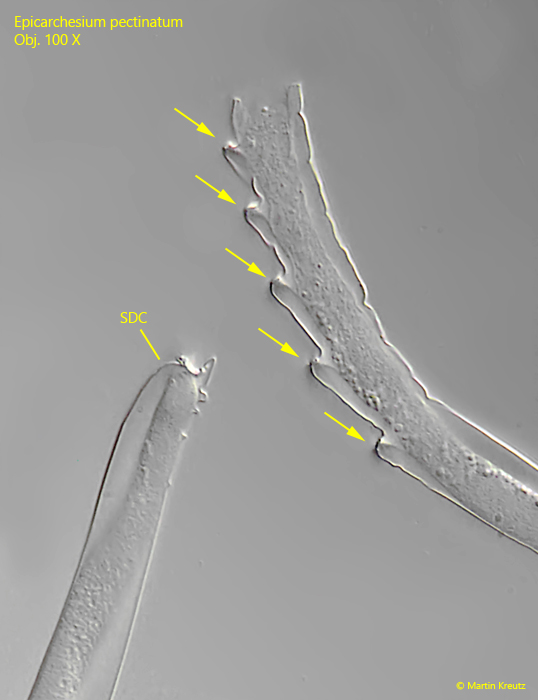
The stalks of the colonies are branched and contracts spirally. Inside the stalks, the string-like myoneme is clearly recognizable (s. fig. 8). At the ramification sites, however, the myoneme is interrupted and provided with a transverse wall. This transverse wall acts like a predetermined breaking point. The daughter cells formed can easily detach from this point. What remains is a wedge-shaped notch. If several daughter cells detach one after the other, a serrated structure is formed (s. fig. 13), which is very characteristic of Epicarchesium pectinatum.
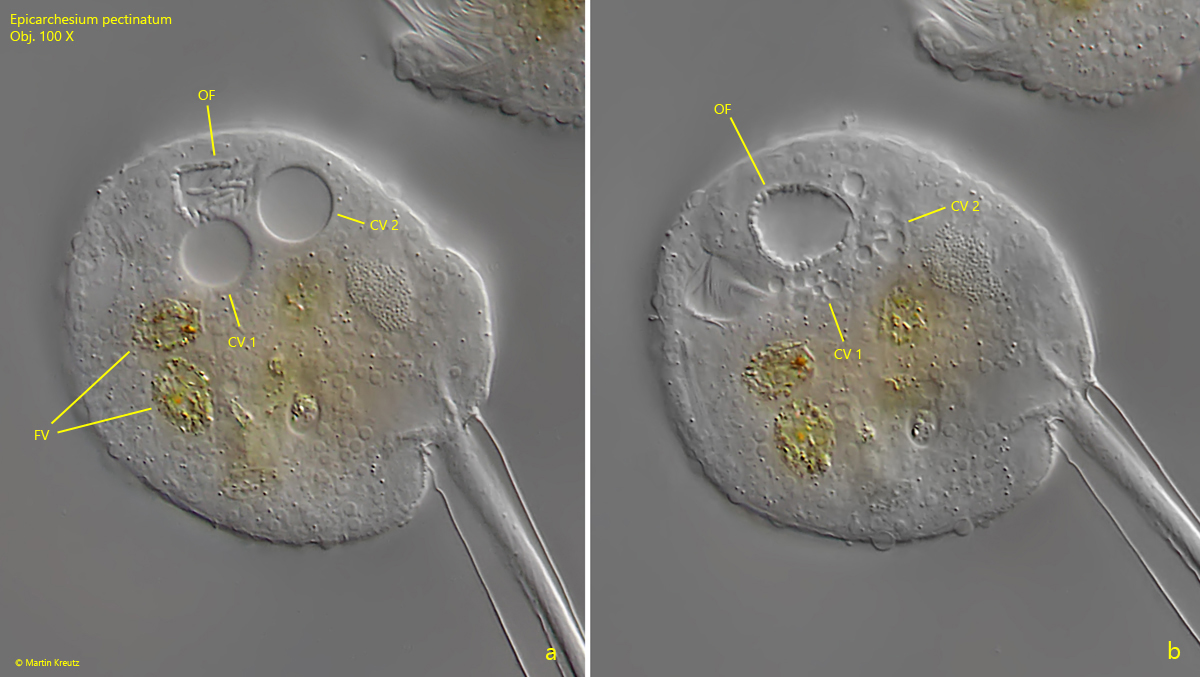
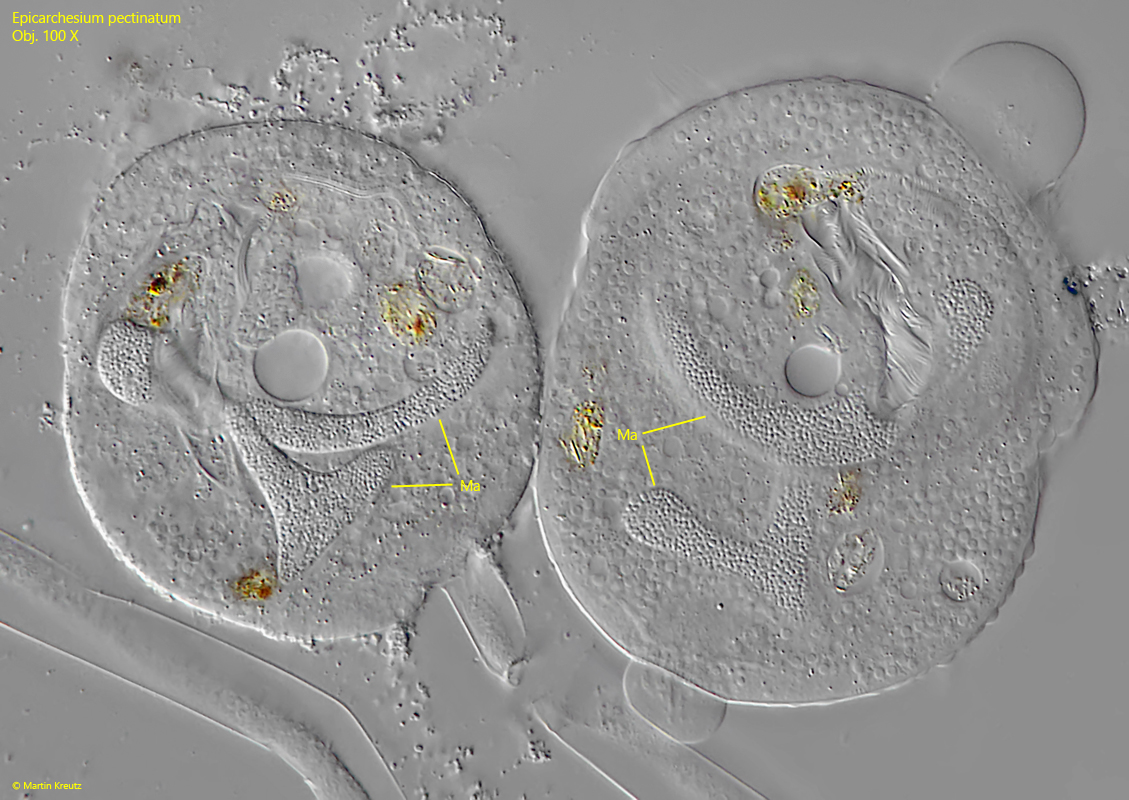
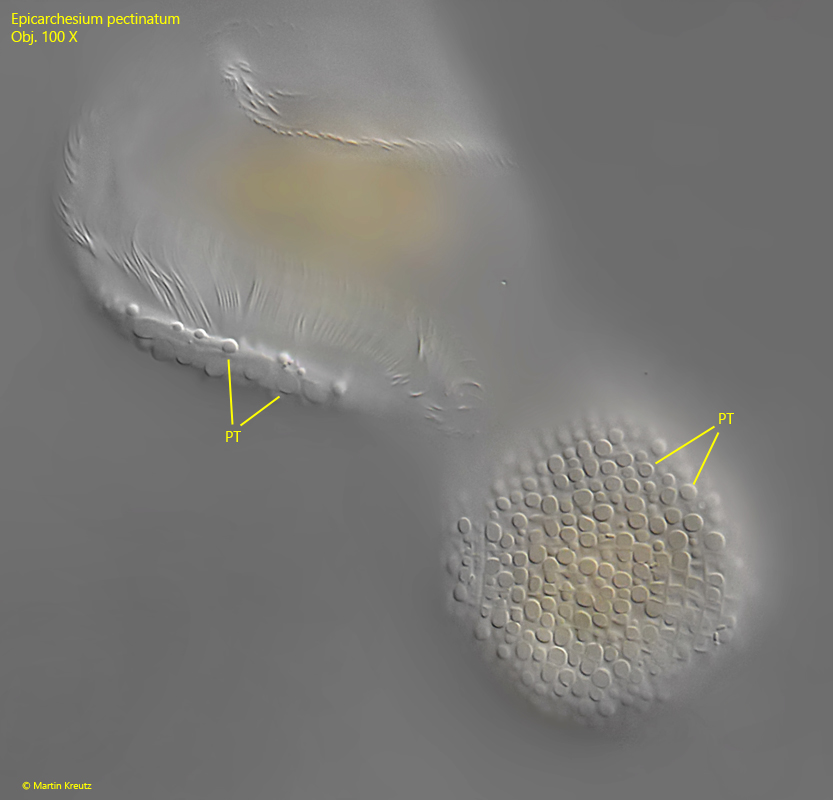
The pellicle and the edge of the peristome are densely covered with tubercles (s. fig. 6) which vary in size and are arranged in transverse rows. The two contractile vacuoles, which empty into the oral funnel, can only be recognized with certainty in slightly squashed specimens (s. fig. 3 a-b). This may be the reason why they were partly overlooked by earlier authors. The macronucleus lies along the longitudinal axis of the cell and is J-shaped (s. fig. 5). I could not identify the micronucleus. However, it is also not mentioned by earlier authors, which is probably why it is difficult to identify.
Fig. 1:Epicarchesium pectinatum. L = 63–68 µm (of zooids). A freely swimming colony with five zooids. Note the swimming direction (arrow). Obj. 20 X.
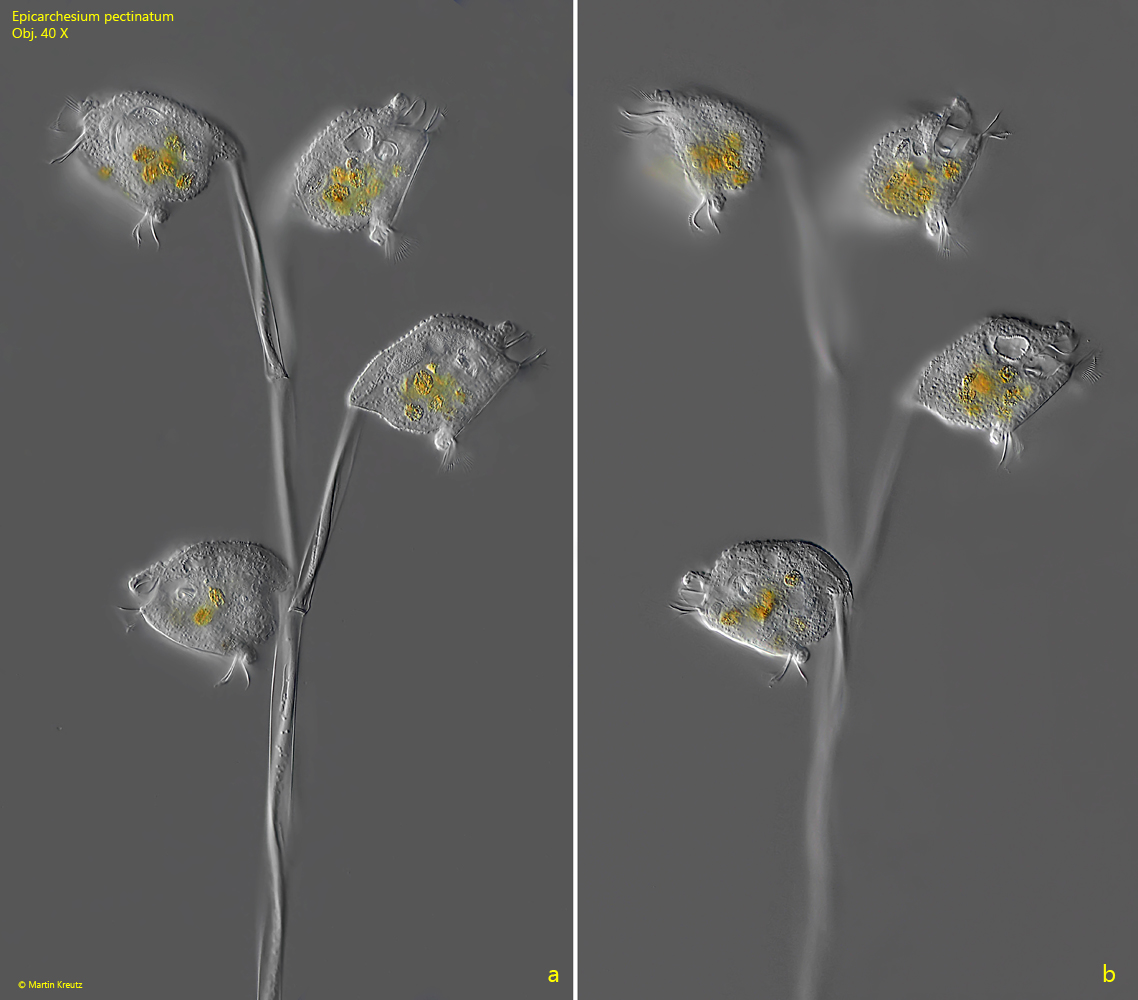
Fig. 2 a-b:Epicarchesium pectinatum. L = 63–68 µm (of zooids). Two focal planes of the colony as shown in fig. 1. Obj. 40 X.
Fig. 3 a-b:Epicarchesium pectinatum. The two contractile vacuoles (CV 1, CV 2) during the diastole (a) and the systole (b). Both contractile vacuoles are empty into the oral funnel (OF) visible in a cross-section. As the two contractile vacuoles empty, the funnel expands due to the inflowing water (b). The inner wall of the oral funnel is covered with ingested bacteria. FV = food vacuoles. Obj. 100 X.
Fig. 4 a-b:Epicarchesium pectinatum. The mouth opening (MO) and the oral funnel (OF) in two specimens. Obj. 100 X.
Fig. 5:Epicarchesium pectinatum. The J-shaped macronuclei (Ma) in two squashed specimens. Obj. 100 X.
Fig. 6:Epicarchesium pectinatum. The pellicle is covered with tubercles (PT) of different size, arranged in transversal rows. Obj. 100 X.
Fig. 7:Epicarchesium pectinatum. The fibers of the myoneme (MY) projecting into the cytoplasm of the zooid. Obj. 100 X.
Fig. 8:Epicarchesium pectinatum. The stalk and a ramification site (RS) in detail. Note the separate myonema in both branches (SM). They are not connected but separated by a transverse wall. The myonema are covered with mitochondria (Mit). Obj. 100 X.
The stalks of Epicarchesium pectinatum have serrated structures at the distal ends in older colonies, which results from the detachment of daughter cells from the colony. To facilitate detachment, the myonema are not connected to each other at the ramification sites. In addition, a transverse wall is formed, which acts as a kind of “predetermined breaking point”. I was able to observe how this serrated structure develops in the colonies of my population.
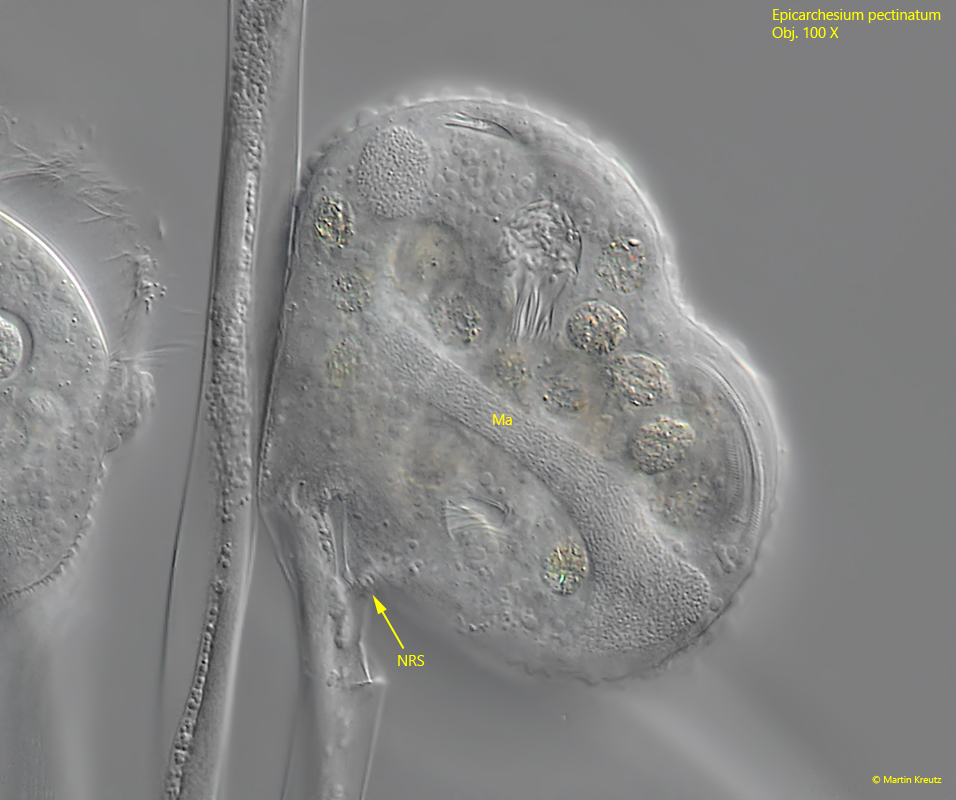
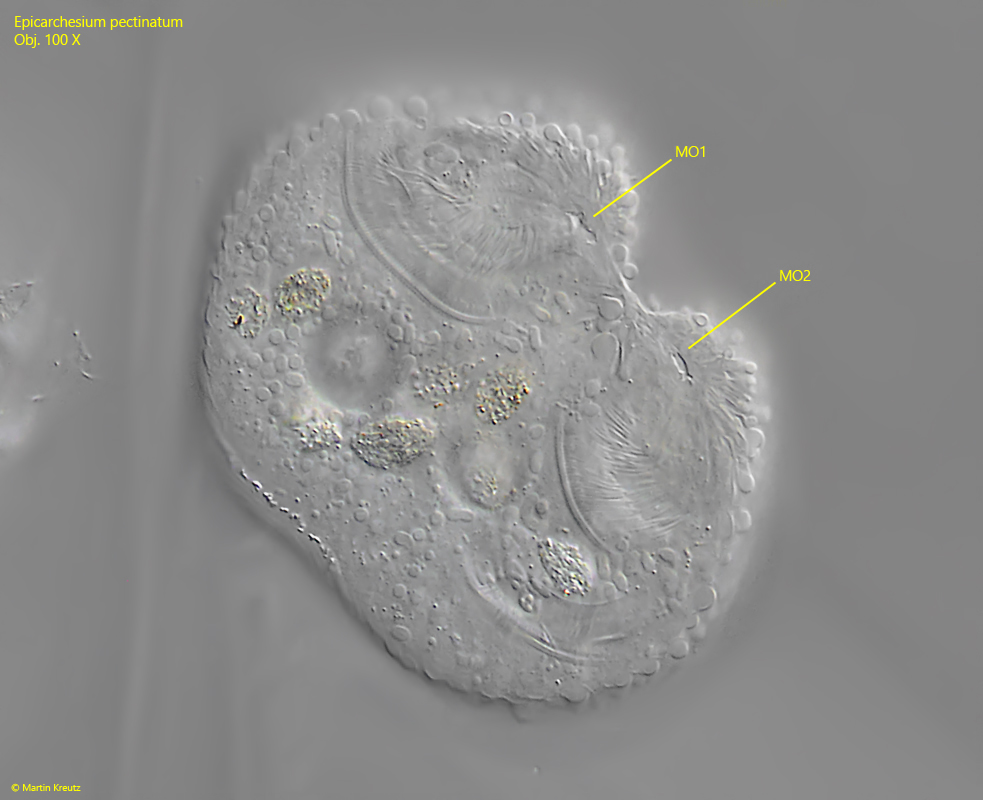
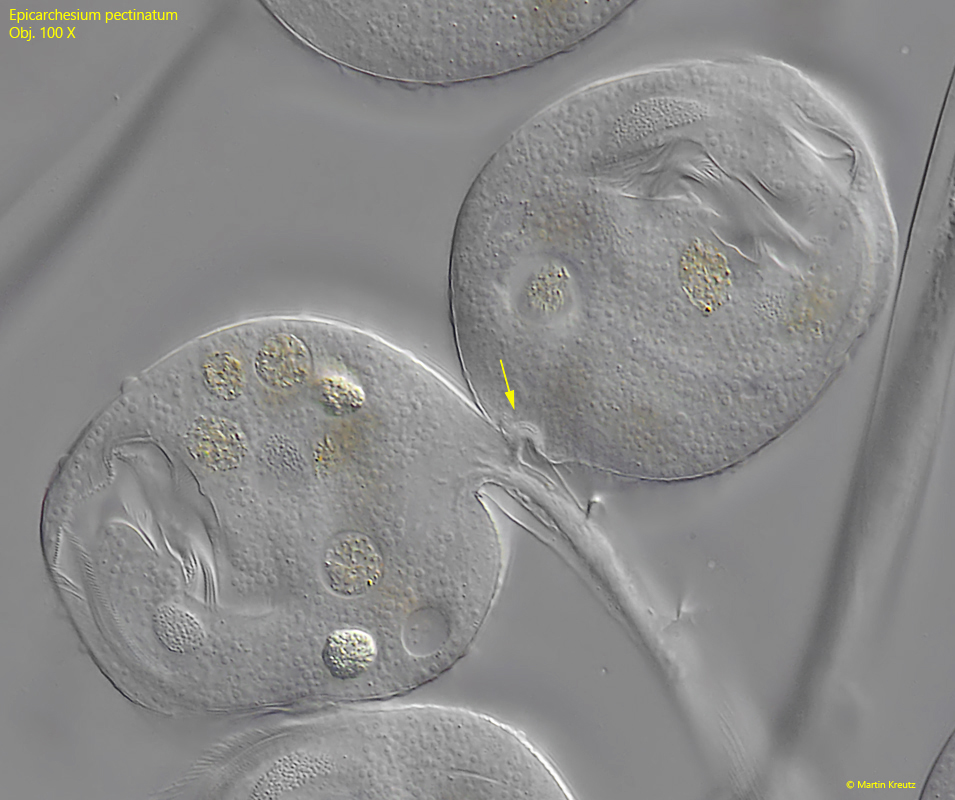
It begins with a cell division of a zooid, which in peritrichous ciliates always takes place along the longitudinal axis (s figs. 9, 10, 11 and 12). During cell division, the stalk-forming organelle (the scopula) is also duplicated and passed on to the daughter cell (s. fig. 9). As a result, the stalk is also duplicated. After the daughter cell has separated from the mother cell, it sits on its own stalk. However, this does not yet have a myoneme (s. fig. 11). A transverse wall is now drawn in at the junction (s. fig. 12). The daughter cell then extends its own stalk with a separate myoneme in it. The longer the stalk of the daughter cell becomes, the greater the mechanical stress on the ramification site, which ultimately leads to the detachment of the daughter cell, which founds a new colony. What remains is a wedge-shaped notch on the stalk (s. fig. 13).
Fig. 9:Epicarchesium pectinatum. A zooid during cell division along the longitudinal axis. The macronucleus (Ma) is just divided. Note the new ramification site (NRS) formed during the cell division. Obj. 100 X.
Fig. 10:Epicarchesium pectinatum. The same specimens as shown in fig. 9 with focal plane on the mouth openings (MO 1, MO 2). Obj. 100 X.
Fig. 11:Epicarchesium pectinatum. The new ramification site (arrow) in the just separated cells. The transverse wall is not yet formed in this phase. Obj. 100 X.
Fig. 12:Epicarchesium pectinatum. Shortly after the completed cell division the transverse wall (TW) is formed at the ramification site. Obj. 100 X.
Fig. 13:Epicarchesium pectinatum. The stalk of a daugther cell (SDC) was detached and has left a wedge-shaped notch on the stem of the colony. By repeating this process, a serrated structure is created (arrows). Obj. 100 X.