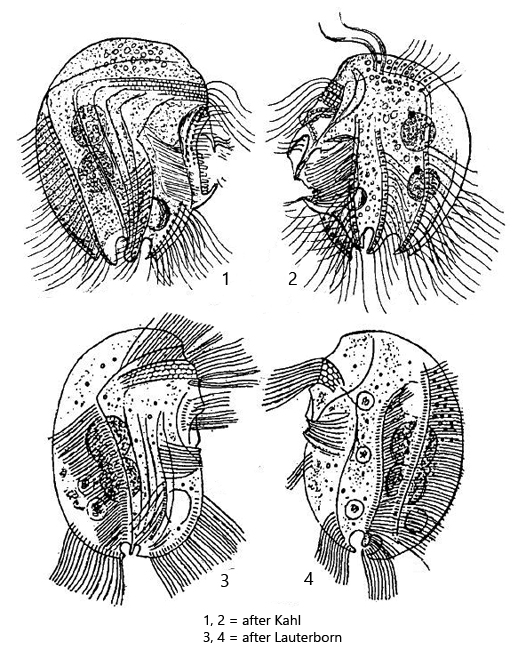
ventral side mostly straight and with ribbed and spine-like projections
dorsal side semicircular, smooth
somatic cilia long and dense, present in 8 longitudinal but somewhat oblique rows. posterior 2 roundish indentations (ω-shaped) surrounded by 6 inconspicuous spines
macronucleus in one or two parts but never more, each with an adjacent micronucleus
contractile vacuole below adoral zone of membranelles
in cytoplasm sometimes endosymbiotic bacteria present
Pelodinium reniforme
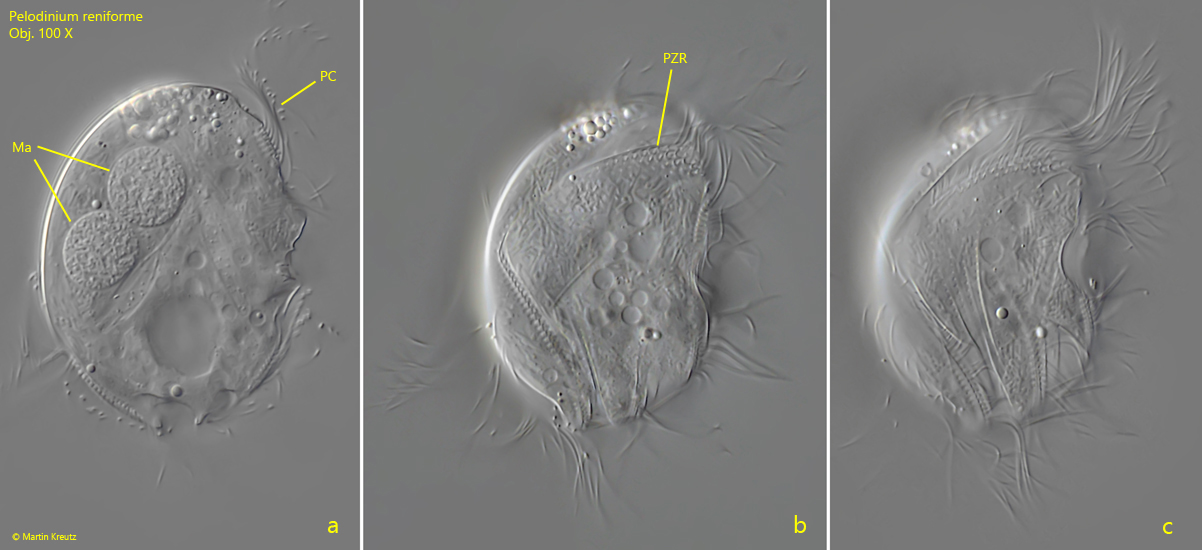
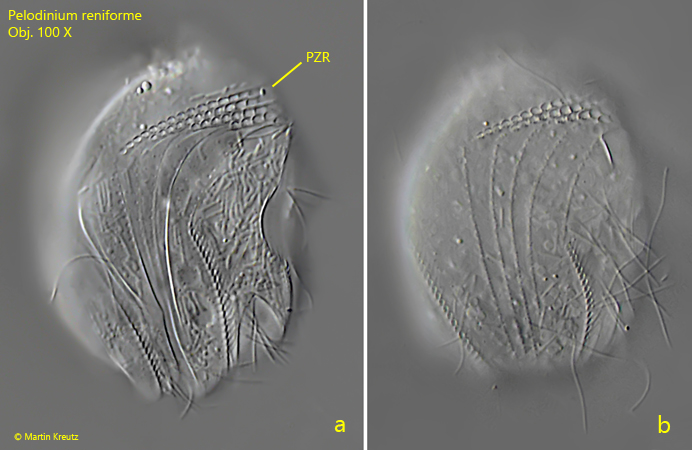
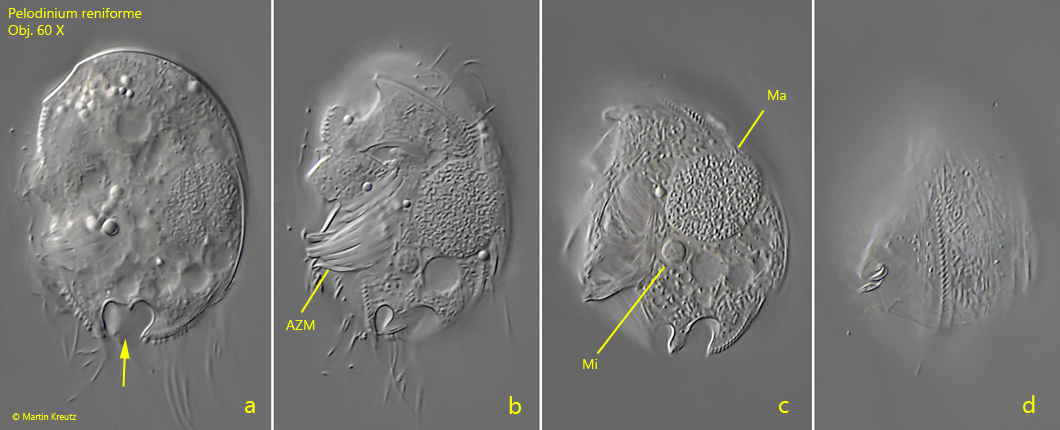
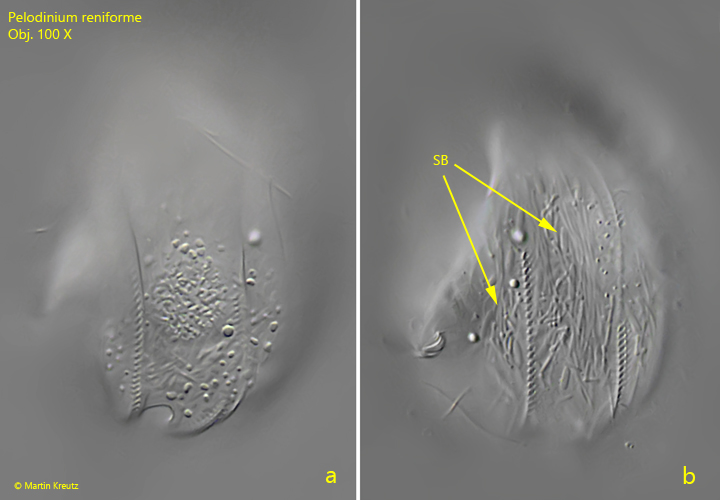
I have found Pelodinium reniforme rarely, but regularly in the mud samples from Simmelried as well as from Purren pond. The species is often associated with other odontostomatid ciliates. The genus Pelodinium has no conspicuous spines or tooth-shaped extensions. The body shape is mostly oval in my population. The somatic ciliation of Pelodinium is long and dense, which is why this species can swim comparatively fast. Under coverslip pressure, the cilia are quickly shed off. However, in rare cases photographs under coverslip pressure with the complete ciliation retained are possible (s. fig. 1 a-d and fig. 2 a-c). Pelodinium reniforme is quite easily recognized by two round notches at the posterior end, which looks like an ω (greek omega) when focused on the middle body axis. In my population I have found specimens with one and two macronuclei. However, both forms are covered by the definition of the species. I could also identify symbiotic bacteria in some specimens (s. fig. 6b), but did not examine them more closely.
Fig. 1 a-c:Pelodinium reniforme. L = 49 µm. Three focal planes of a slightly squashed specimen from right. Ma = macronuclei, PC = perizonal row of cilia, PZR = perizonal row. Obj. 100 X.
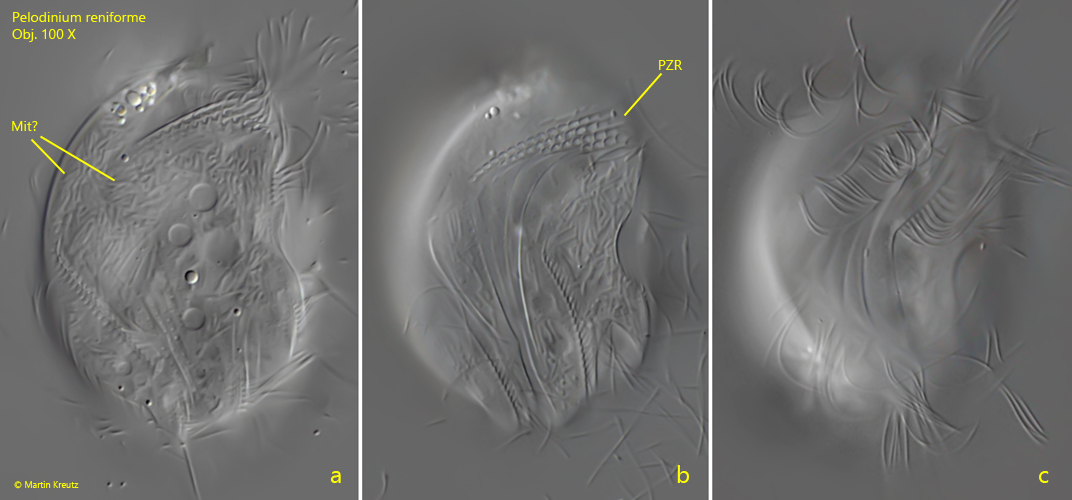
Fig. 2 a-c:Pelodinium reniforme. L = 49 µm. A more detailed view on the right side of the same specimen as in fig. 1 a-c. Mit? = most probably mitochondria below the pellicle (symbiotic bacteria are not curved and rod-shaped), PZR = perizonal row. Obj. 100 X.
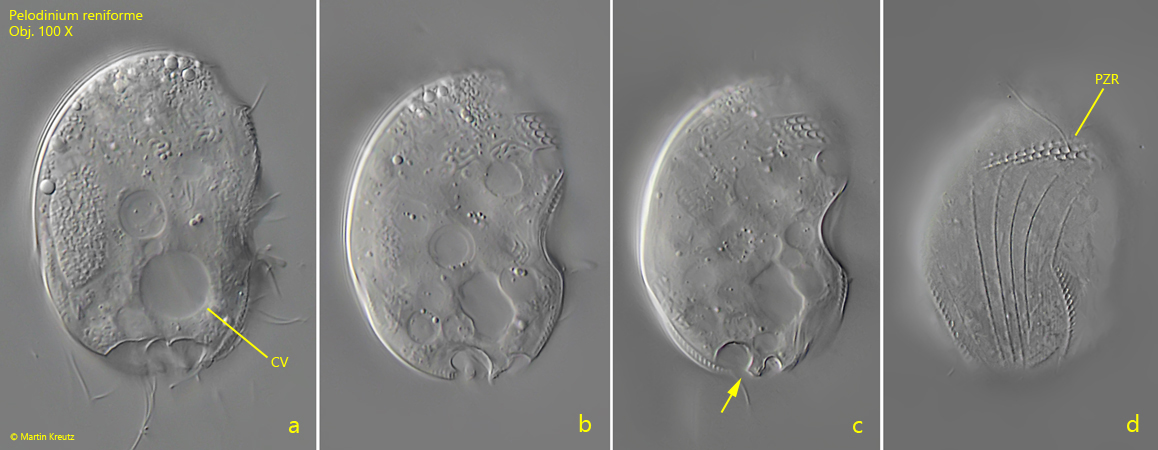
Fig. 3 a-d:Pelodinium reniforme. L = 46 µm. Four focal planes from right of a slightly squashed, second specimen. Note the ω-shaped indentations at the posterior end (fig. 2c, arrow). CV = contractile vacuole, PZR = perizonal row. Obj. 100 X.
Fig. 4 a-b:Pelodinium reniforme. The structure of the right side of a more strongly squashed specimen in detail. PZR = perizonal row. Obj. 100 X.
After transferring a sample to the slide, the specimens of Pelodinium reniforme preferentially settle down on the left side of the body, which is why in most cases the right side is turned to the coverslip. The left side of the body, which is structured in a much more inconspicuous way, can be examined much less frequently (s. fig. 5 a-d and fig. 6 a-b).
Fig. 5 a-d:Pelodinium reniforme. L = 43 µm. Four focal planes from left side of a slightly squashed specimen. Note the ω-shaped indentations at the posterior end (fig. 4a, arrow). AZM = adoral zone of membranelles, Ma = macronucleus, Mi = micronucleus. Obj. 60 X. Obj. 100 X.
Fig. 6 a-b:Pelodinium reniforme. The structure of the left side in detail at higher magnification. In this specimen symbiotic bacteria (SB) are visible. Obj. 100 X.
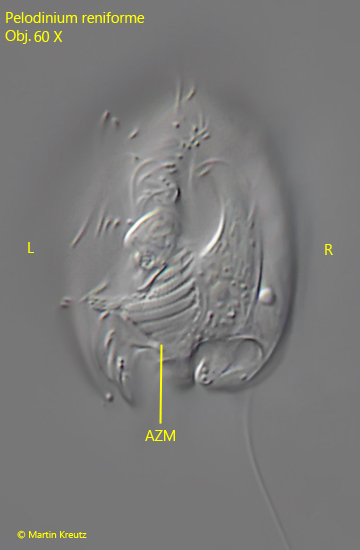
The ventral side of Pelodinium reniforme with the adoral zone of membranelles is only visible in freely swimming specimens. In rare cases, specimens attach themselves to the coverslip with the ventral side for a few seconds. The photo in fig. 7 was taken on such an occasion.
Fig. 7:Pelodinium reniforme. L = 49 µm. The ventral side of a freely swimming specimen. AZM = adoral zone of membranelles, L = left side, R = right side. Obj. 60 X.