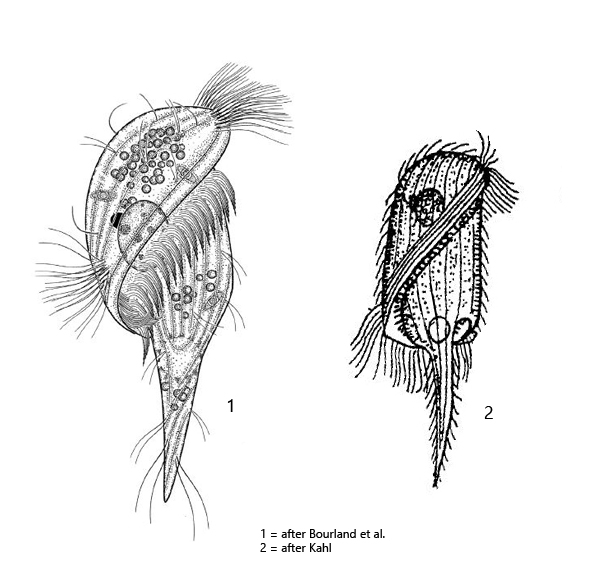
body slenderly oval to oval with a tail-shaped posterior end
dorso-ventrally somewhat flattened
length about 80–130 µm
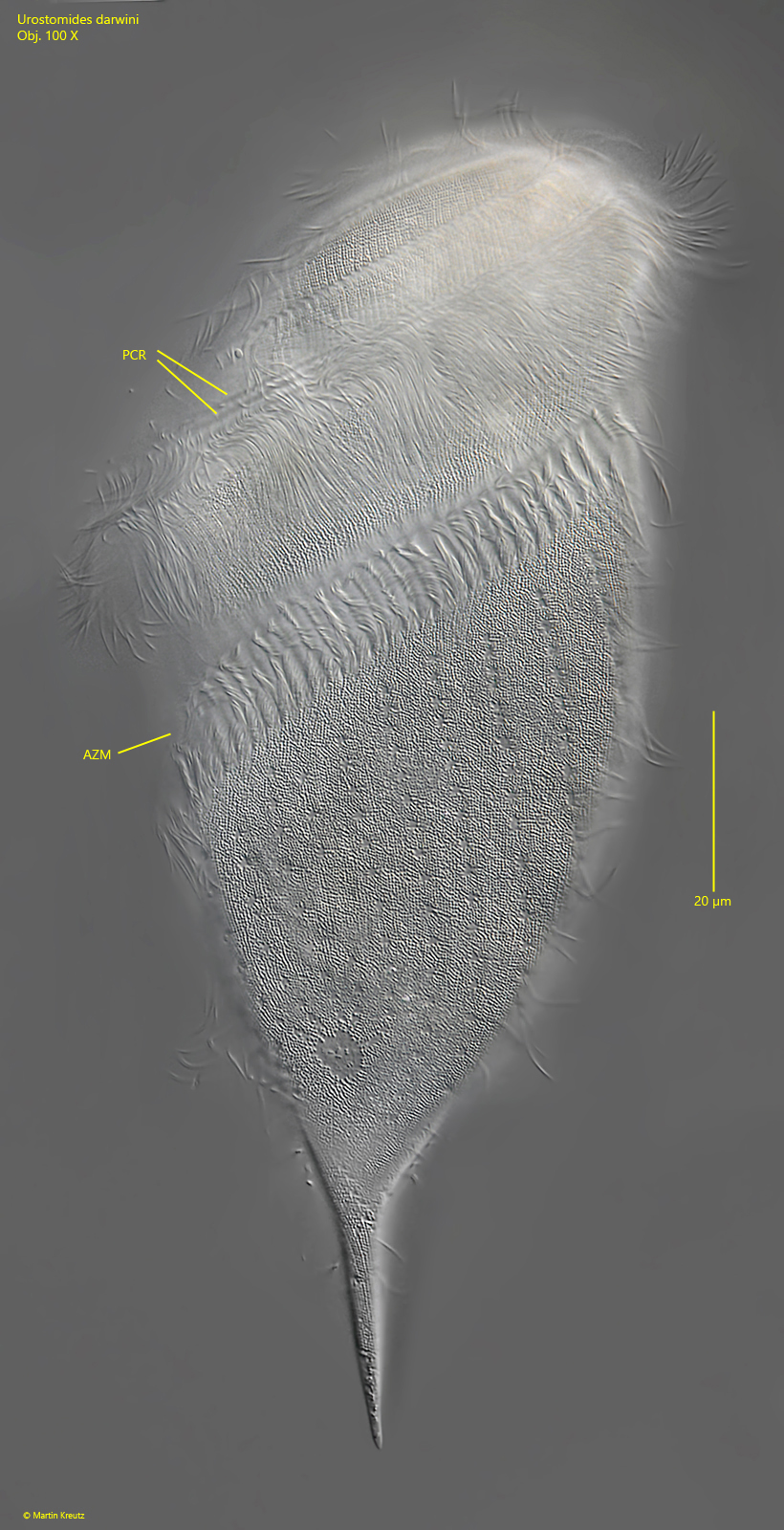
adoral zone running a full turn around the longitudinal axis
adoral zone is covered by the dome towards the posterior end
a small membranella in front of the mouth funnel a small membranelle
periozonal zone consists of 5 rows
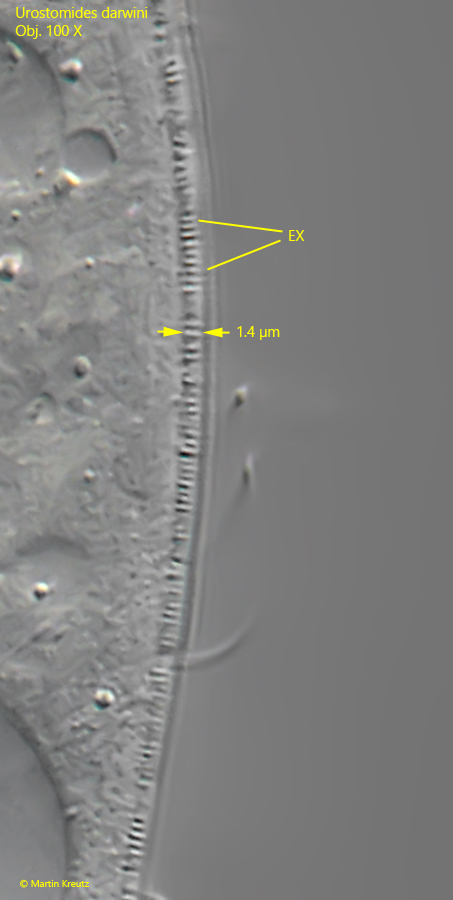
a fringe of extrusomes beneath the pellicle
whole body ciliated, also the caudal spine
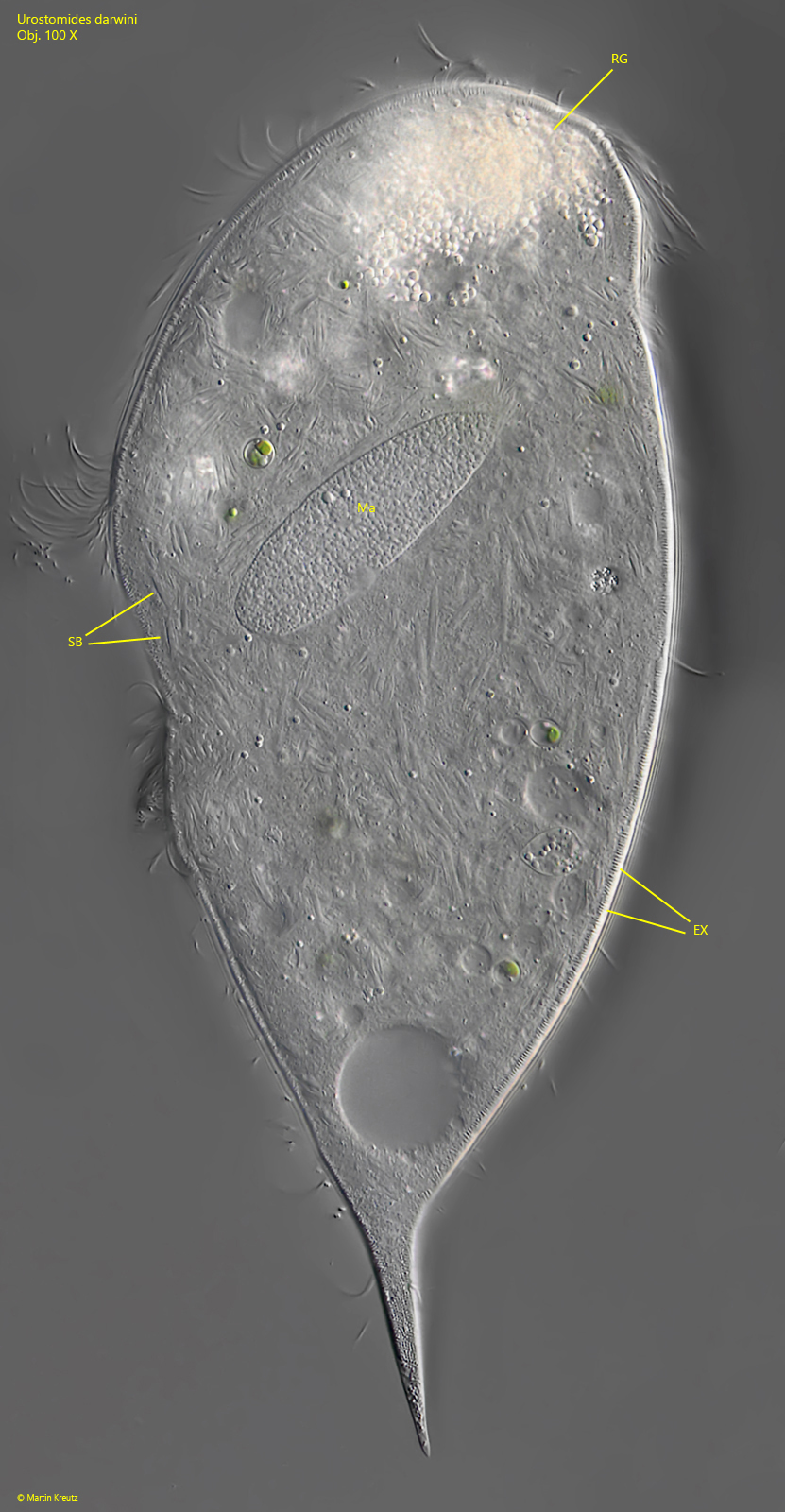
macronucleus globular
contractile vacuole at the base of the caudal spine
Urostomides darwini
Urostomides darwini was first described as Metopus darwini by Kahl in 1927. Later Jankowski (1964) transferred the species to the genus Brachonella. For the genus Brachonella, Jankowski defines the following characteristics:
– anterior end (dome) larger than the posterior end. – the adoral zone forms a helical line when it runs to the mouth opening
In 2017 Bourland et al. provide a redescription and 18S rRNA gene sequences. Based on these results Brachonella (Metopus) darwini was transferred to the genus Urostomides.
Regarding the occurence of Urostomides darwini Kahl writes: “stets nur vereinzelt, aber an vielen Fundstellen (always only sporadically, but in many localities)”. I also find Urostomides darwini in many localities. The population densities vary a lot. Sometimes I find no specimens for 1–2 years and sometimes up to 10 specimens per milliliter. The reasons for these fluctuations are not known to me.
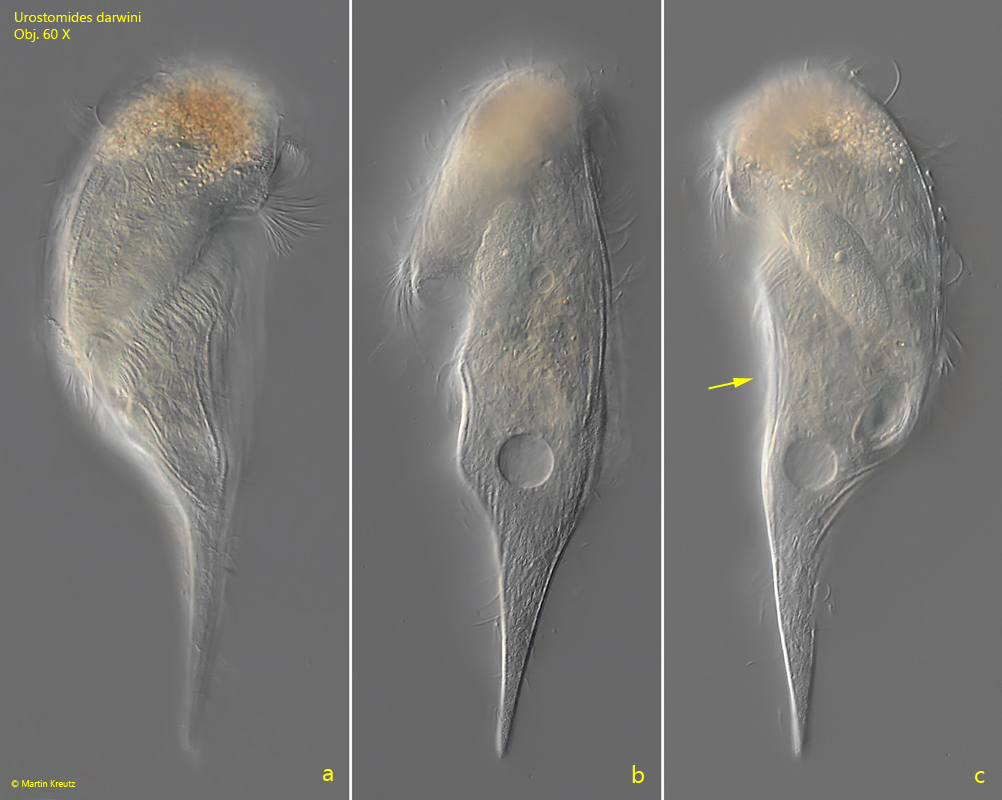
At high layer thickness Urostomides darwini can be observed quite well even with the coverslip (s. fig. 1 a-c). It swims quite fast and rotates around its own axis. The specimens appears mushroom shaped with a crooked cap (s. fig. 1 a-c). In the freely swimming specimen I could see that the right side is concavely depressed (s. fig. 1 c), which Kahl did not mention and drew. The specimens in my population were mostly yellowish in color, which differs from Kahl’s description, who describes the cytoplasm as colorless. The specimens in my population were also significantly larger than described by Kahl and Bourland et al. with a length of 150–200 µm. Thus, they were about 30% larger than indicated by these authors.
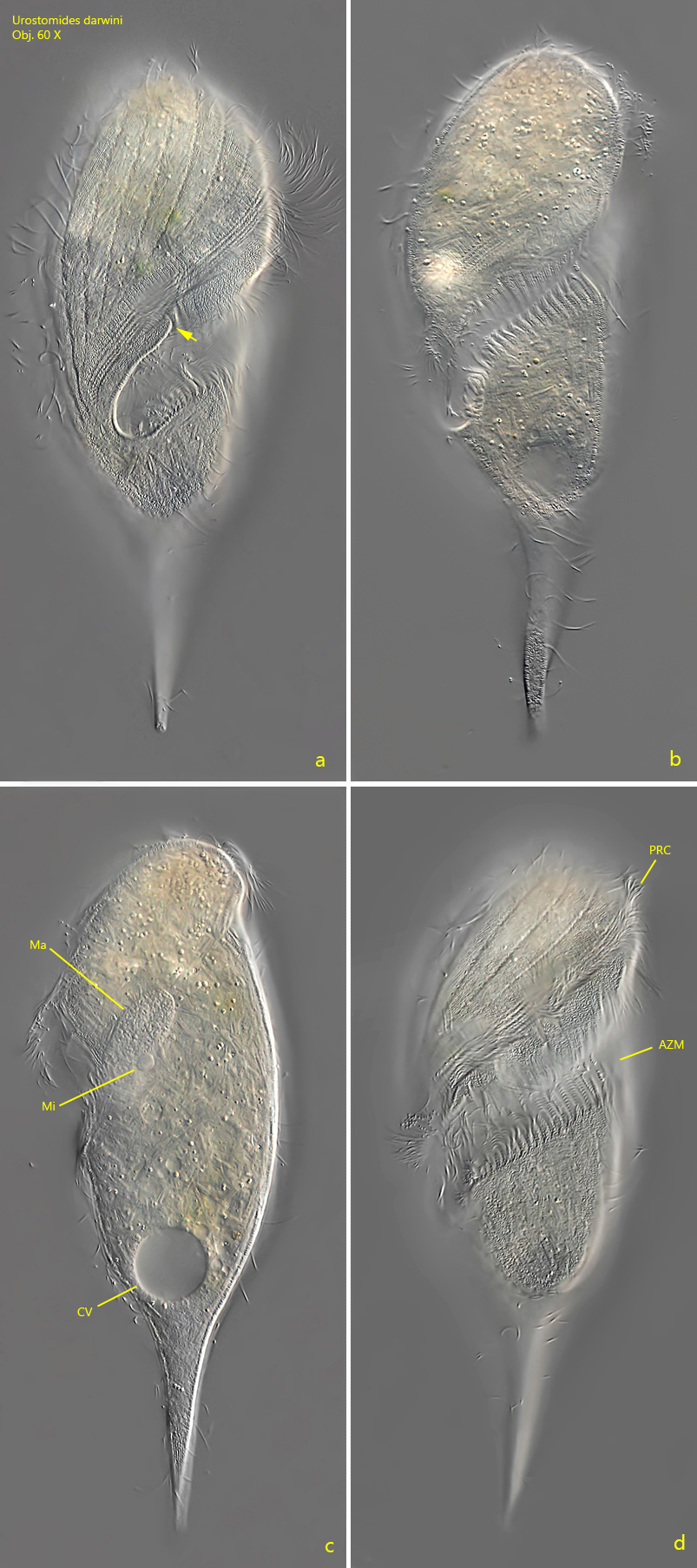
Kahl considered Urostomides darwini as a transitional form to the genus Caenomorpha, which can be easily understood because of the body shape with the caudal spine. The adoral zone crosses the ventral side diagonally (s. fig. 1 d) and continues to the right side where the mouth opening is located (s. fig. 1 a). On the right side the adoral zone descends into the mouth funnel. At the point where it descends, a cytoplasm lip forms the outer edge of the mouth opening (s. fig. 2 a). The macronucleus was always oval in my specimens (Kahl and Bourland et al. describe it as globular). The spherical micronucleus is adjacent to the macronucleus and has a diameter of 5–6 µm (s. fig. 2 c). The food vacuoles I could find mainly contained rhodobacteria, bacteria, and small algae. Like many other metopid ciliates, Urostomides darwini also bears an aggregation of refractive, brownish granules of unknown function in the apical dome (s. fig. 5).
Fig. 1 a-c:Urostomides darwini. L = 188 µm. Ventral view (a), from right (b) and from dorsal (c) of a freely swimming specimen. Note the concave shape of the right margin (arrow, c). Obj. 60 X.
Fig. 2 a-d:Urostomides darwini. L = 200 µm. A slightly squashed specimen from right (a, b, c) and ventral (d). Note the cytoplasm lip (arrow, a) where the adoral zone runs into the mouth opening. AZM = adoral zone of membranelles, CV = contractile vacuole, Ma = macronucleus, Mi = micronucleus, PRC = perizonal rows of cilia. Obj. 60 X.
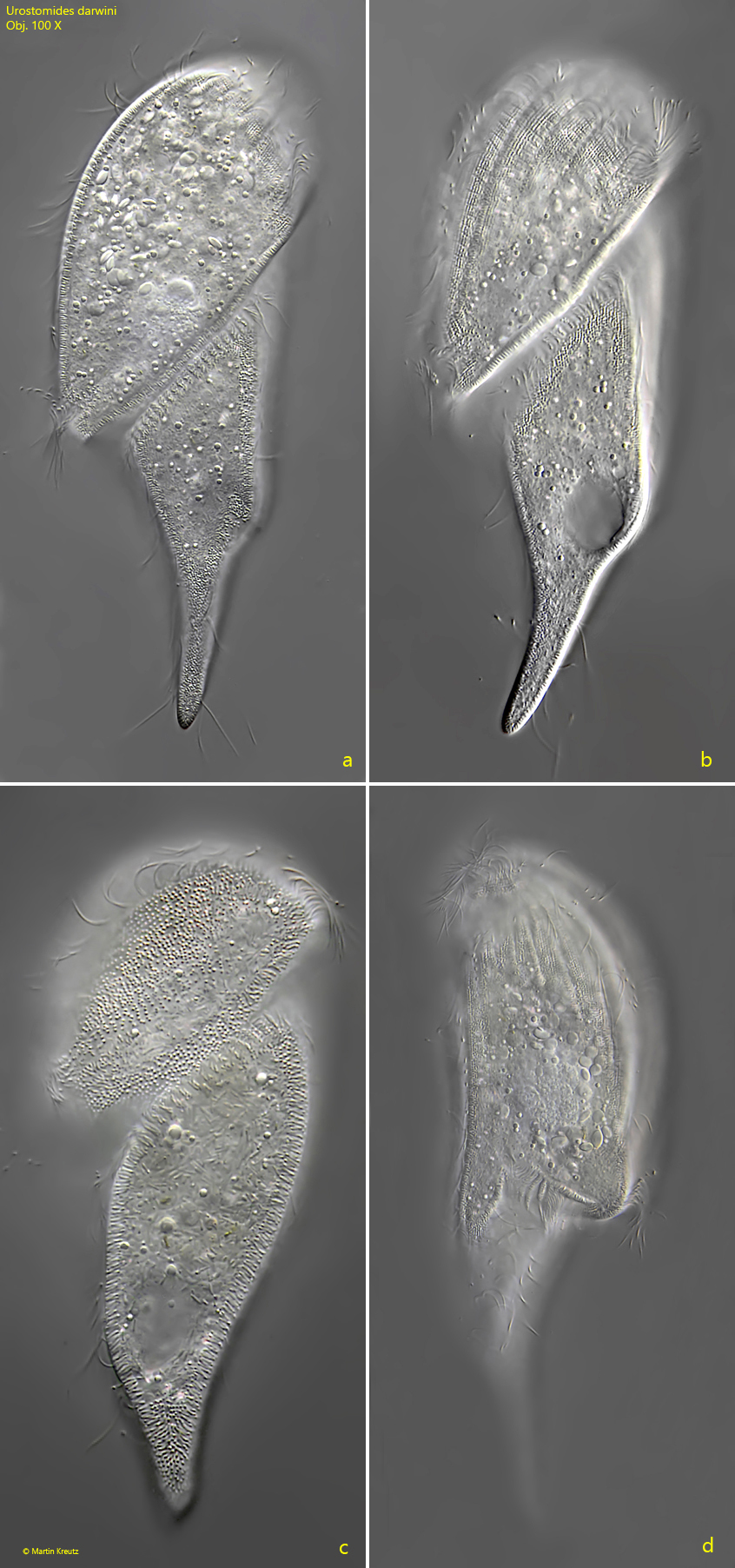
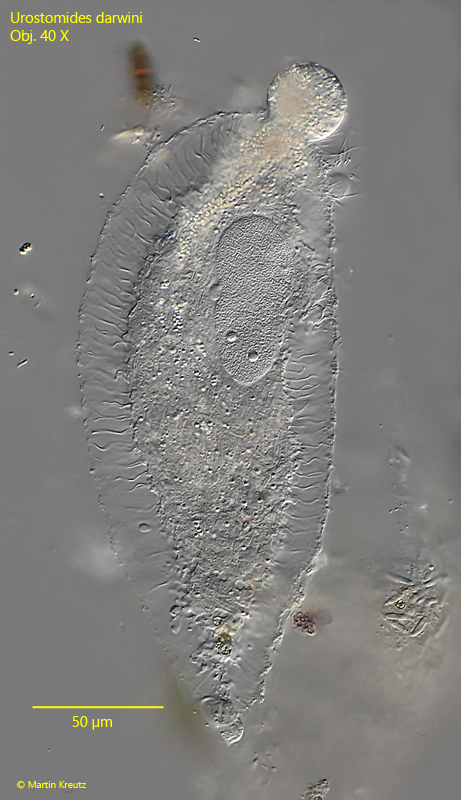
Fig. 3 a-d:Urostomides darwini. L = 150 µm. A second slightly squashed specimen from ventral (a, b, c) and from right (d). Obj. 100 X.
Fig. 4:Urostomides darwini. L = 150 µm. Focal plane on the cell surface of a squashed specimen. AZM = adoral zone of membranelles, PCR = perizonal cilia rows. Obj. 100 X.
Fig. 5:Urostomides darwini. The same specimen shown in fig. 4 but with focal plane on the macronucleus (Ma). Note the large number of symbiotic bacteria (SB) in the cytoplasm of the cell. EX = fringe of extrusomes, RG = aggregation of refractive granules in the apical dome. Obj. 100 X.
Fig. 6:Urostomides darwini. The fringe of extrusomes (EX) beneath the pellicle in detail. In this specimen the rod-shaped extrusomes are 1.4 µm long. Obj. 100 X.
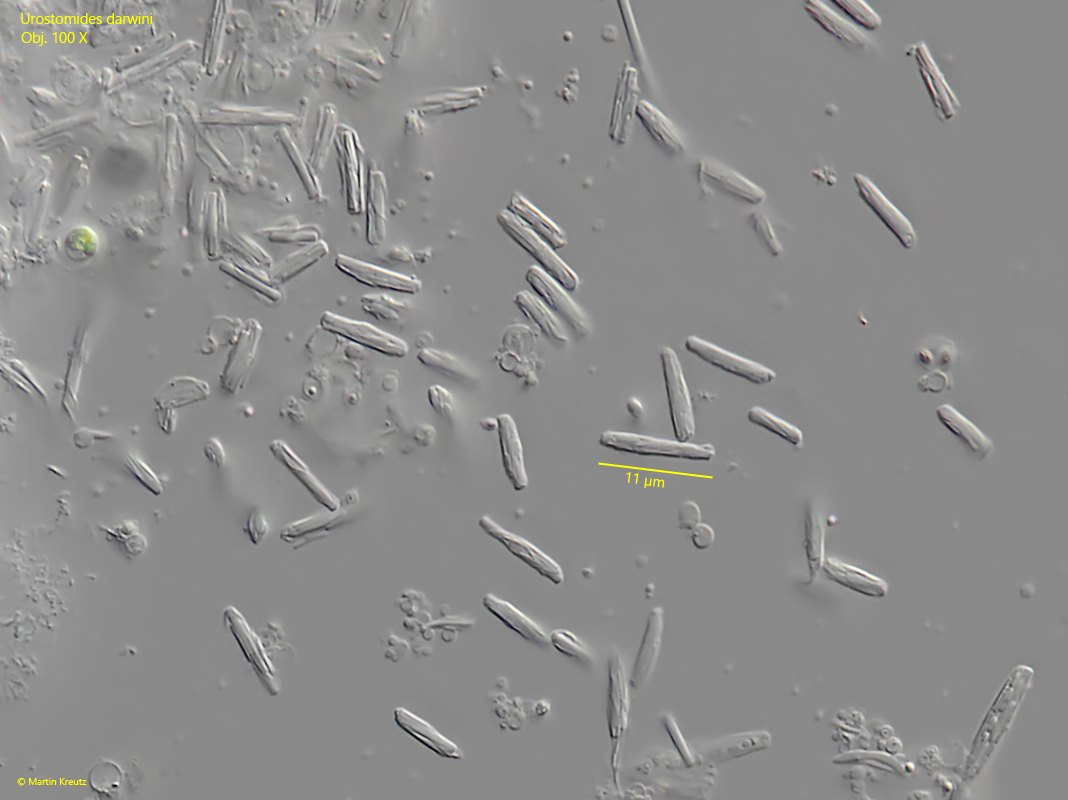
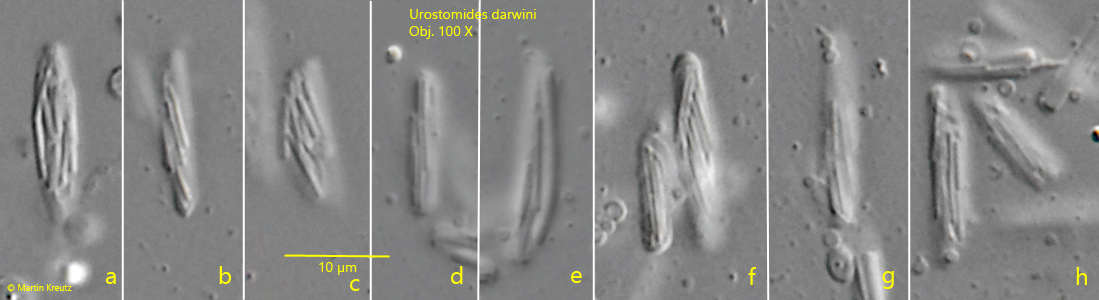
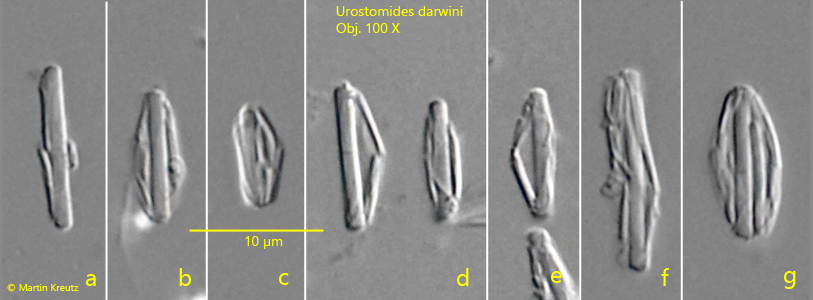
Urostomides darwini does not possess mitochondria, but bacterial symbionts that enable it to live in an anaerobic environment. They are methanogenic bacteria that degrade metabolites of the ciliate to methane. The symbiotic bacteria of Urostomides darwini differ significantly from the symbiotic bacteria I have found so far in other metopid ciliates. They are distributed throughout the cytoplasm and fill virtually the entire ciliate (s. figs. 5 and 7). However, the bacteria are not solitary, but seem to form aggregates. Comparatively large bacteria with a length of about 10 µm are covered by a layer of much smaller bacteria (s. fig. 8 a-h). When the layer thickness is strongly reduced, these smaller bacteria are detached (s. fig. 9 a-g). They seem to adhere firmly to these large bacteria. The large bacteria are also exposed during this procedure. They are smooth and cylindrical with rounded ends (s. fig. 9 a-g). The smaller, detached bacteria are about 2–5 µm and 1 µm thick. These could be functional aggregates (or consortia), in which the smaller, superimposed bacteria further process the metabolites of the large central rods and in turn supply the large rods with their metabolites. The exact nature of these aggregates can only be clarified by further investigations.
Fig. 7:Urostomides darwini. The symbiotic bacteria in a strongly squashed specimen. Obj. 100 X.
Fig. 8 a-h:Urostomides darwini. Several aggregates of symbiotic bacteria in the cytoplasm. The aggregates consist of a central, large bacterium with a length of about 10 µm covered by a layer of thin rods of another type of bacterium. Obj. 100 X.
Fig. 9 a-g:Urostomides darwini. Under the pressure of the coverslip, the thin bacteria covering the central bacterium get detached and reveal the structure of the aggregates. Obj. 100 X.
To obtain high-resolution images of Urostomides darwini, as shown in figs. 4 and 5, specimens must be squashed under the coverslip. However, this almost always fails due to the strong coverglass sensitivity of this species. As a reaction the extrusomes (mucocysts) get ejected with simultaneous shrinkage and denaturation of the cell body and the the formation of a kind of mucous envelope around the ciliate (s. fig. 10). Surprisingly, however, very few specimens do not seem to be coverslip sensitive. The reason for this is unclear to me. Thus, according to my experience many specimens have to be investigated before one specimen is found which does not denature.
Fig. 10:Urostomides darwini. A denatured specimen after the coverslip has been applied. Obj. 40 X