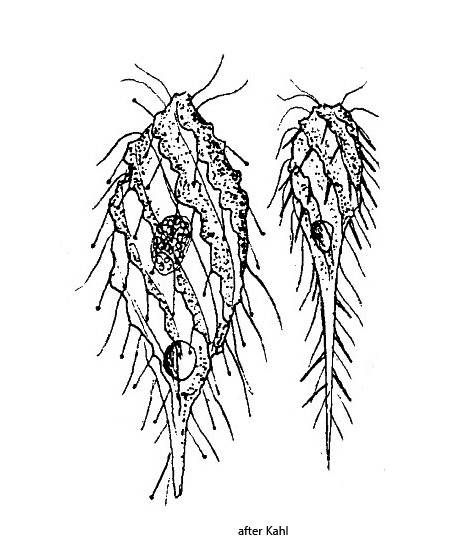
body fusiform to club-shaped, with furrows running counterclockwise in spirals
length 85–110 µm, width 25–35 µm
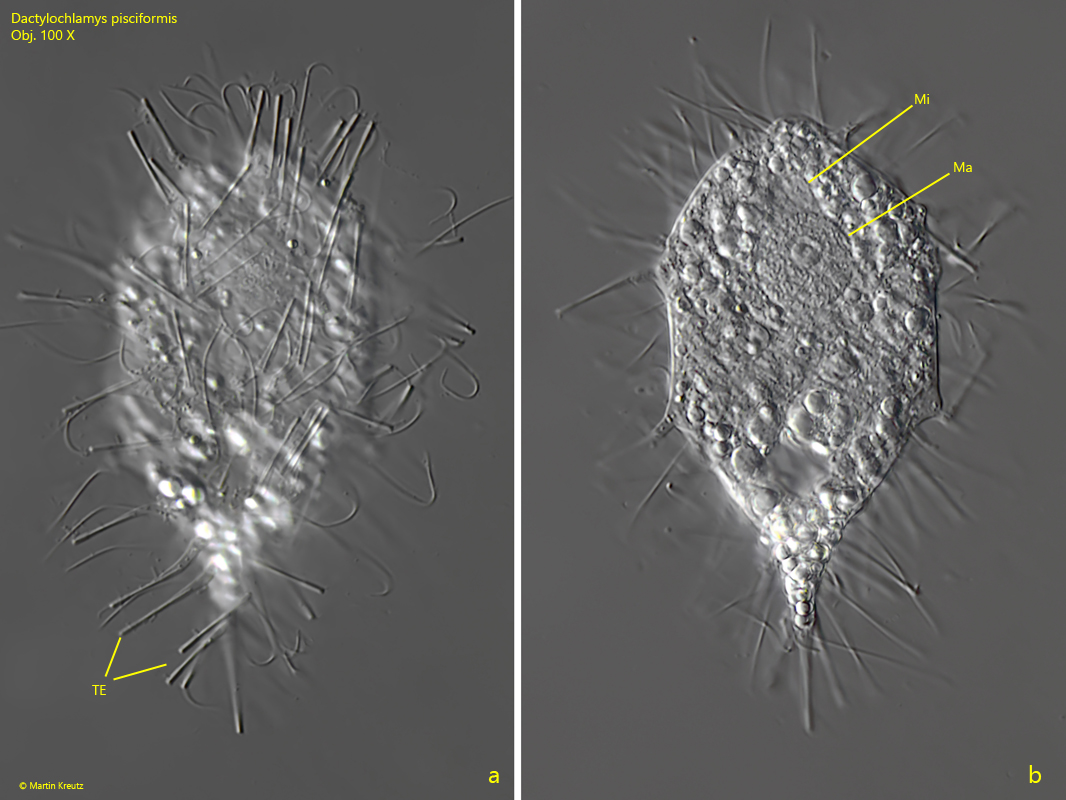
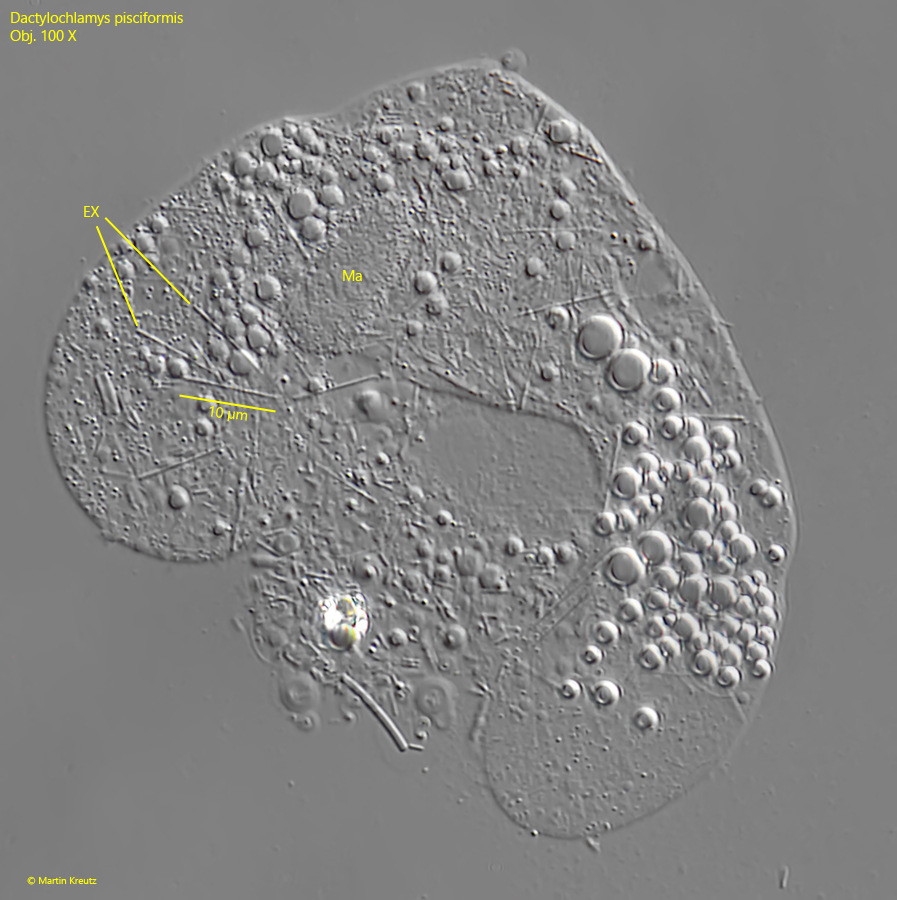
macronucleus globular or ellipsoidal, about 6 x 8 µm
one micronucleus adjacent to macronucleus, diameter about 2–3 µm
many retractable tentacles arise from cell surface
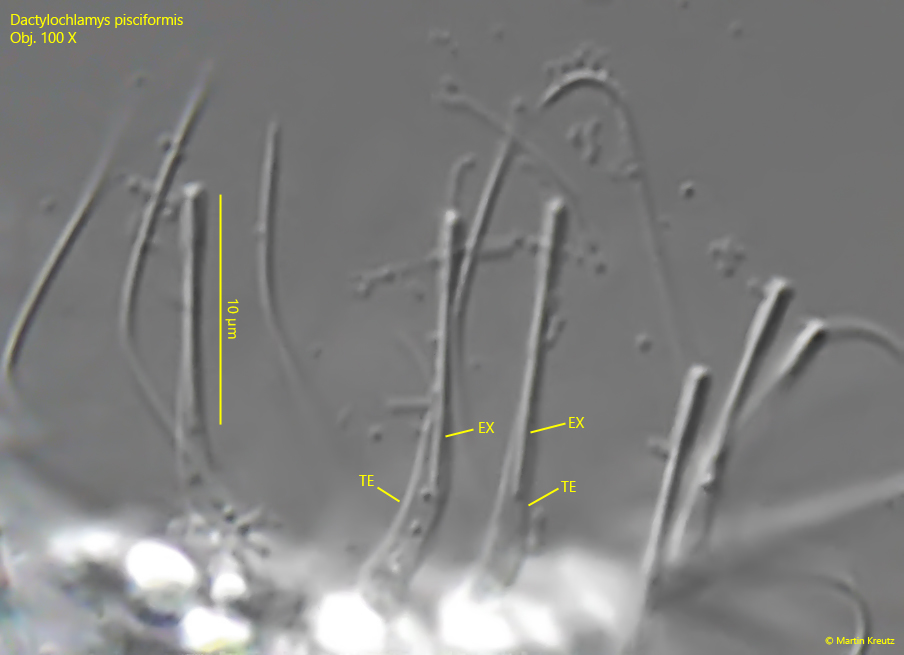
rod-shaped, 10 µm long extrusomes in the distal ends of the tentacles
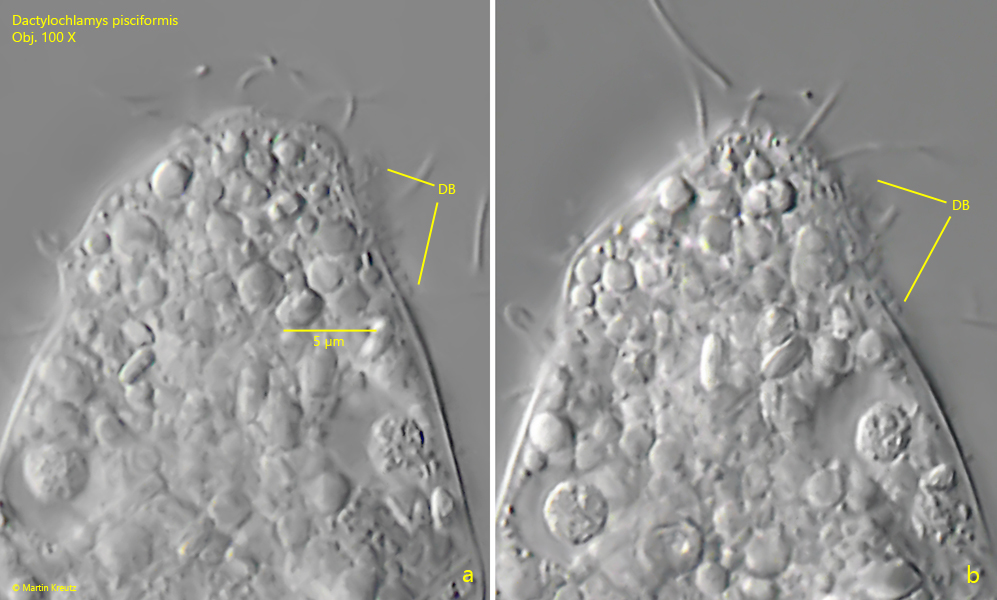
a short dorsal brush (about 5 µm) with short, thickened cilia
cytoplasm filled with refractive globules and food vacuoles
contractile vacuole subterminal
Dactylochlamys pisciformis
I find Dactylochlamys pisciformis regularly in several of my sampling sites, mainly between decaying leaves. In the past years Dactylochlamys pisciformis also occurred at times in high concentrations with about 10 specimens per milliliter. This is also remarkable because the species is described as very rare. When I reported my findings to Foissner in March 2000, he had never found this species before.
There are only a few descriptions of Dactylochlamys pisciformis, mainly by Penard and Kahl. Only recently has Dactylochlamys pisciformis been reexamined microscopically as well as genetically by Pomahač et al. (2023) to determine its phylogenetic position. Indeed, Dactylochlamys pisciformis is a ciliate with curious characteristics. It possesses contractile tentacles, which bear great resemblance to those of the Suctoria or the genus Actinobolina. Therefore, Dactylochlamys pisciformis was originally placed by Kahl in the family Actinobolinidae, which later authors have adopted. According to the analyses of Pomahač et al. Dactylochlamys pisciformis belongs to the spathidiid ciliates and is closely related to the genus Legendrea (e.g. Legendrea loyezae and Legendrea pespelicani).
The specimens of Dactylochlamys pisciformis in my population were very variable in shape, as also described by Kahl. In general, however, the body shape was rather club-shaped. My specimens were between 60–100 µm long and thus slightly smaller than indicated by Kahl and Pomahač et al.
Deviating from the descriptions of Kahl and Pomahač et al., I was able to detect a lamellar cytoplasm lip on the dorsal margin of the mouth bulge in many (but not all) specimens in my population. This cytoplasm lip appeared to be a constant feature and not an artifact (s. figs. 2 a, 3 a and 3 c). In addition, I was able to identify the dorsal brush, which previous authors did not mention (s. fig. 10 a-b). This may be due to the fact that it is very difficult to observe because it is located in a furrow behind the oral bulge and is also very inconspicuous. According to my measurements it is only 5–7 µm long. The thickened special cilia are only 0.7–1 µm long. The presence of a dorsal brush strengthens the affiliation to the spathidiid ciliates.
To my knowledge the structure of the retractable tentacles of Dactylochlamys pisciformis has not yet been studied in detail yet. Although I have been able to observe over 100 specimens, I have unfortunately never been able to find a specimen with fully extended tentacles. However, I found several specimens with partially extended tentacles (s. fig. 4 a-b). In these specimens I could observe that one of the 10 µm long rod-shaped extrusomes is placed in each of the tentacles. The distal ends of the tentacles are cylindrically thickened, but not by the extrusome they carry. The thickening must result from some other structure. At the base, the tentacles are accordion folded when not extended, as is the case with Suktoria. I could also observe that there are “loaded” and “unloaded” tentacles. Some contained an extrusome and others were empty. Possibly these empty tentacles had had contact with a prey organism shortly before. Nothing is currently known about the prey of Dactylochlamys pisciformis. However, I could find paramylon grains in some specimens. Possibly euglenid flagellates are a potential prey.
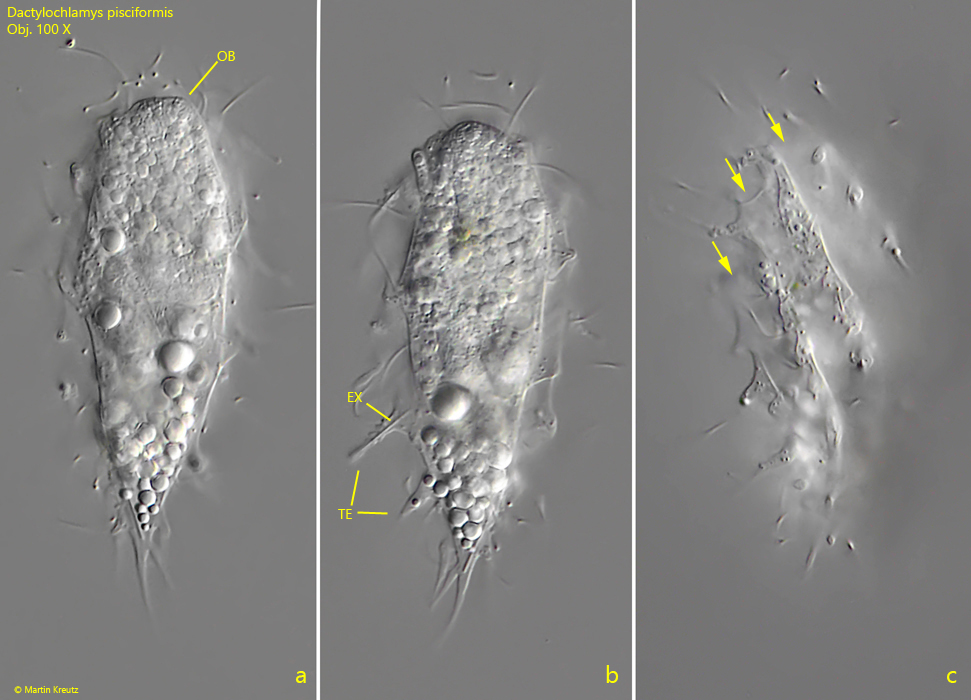
Fig. 1 a-c: Dactylochlamys pisciformis. L = 68 µm. Three focal planes of a freely swimming specimen. Note the spirally running furrows of the body (arrows) and the rod-shaped extrusome in the tentacle (TE). OB = oral bulge. Obj. 100 X.
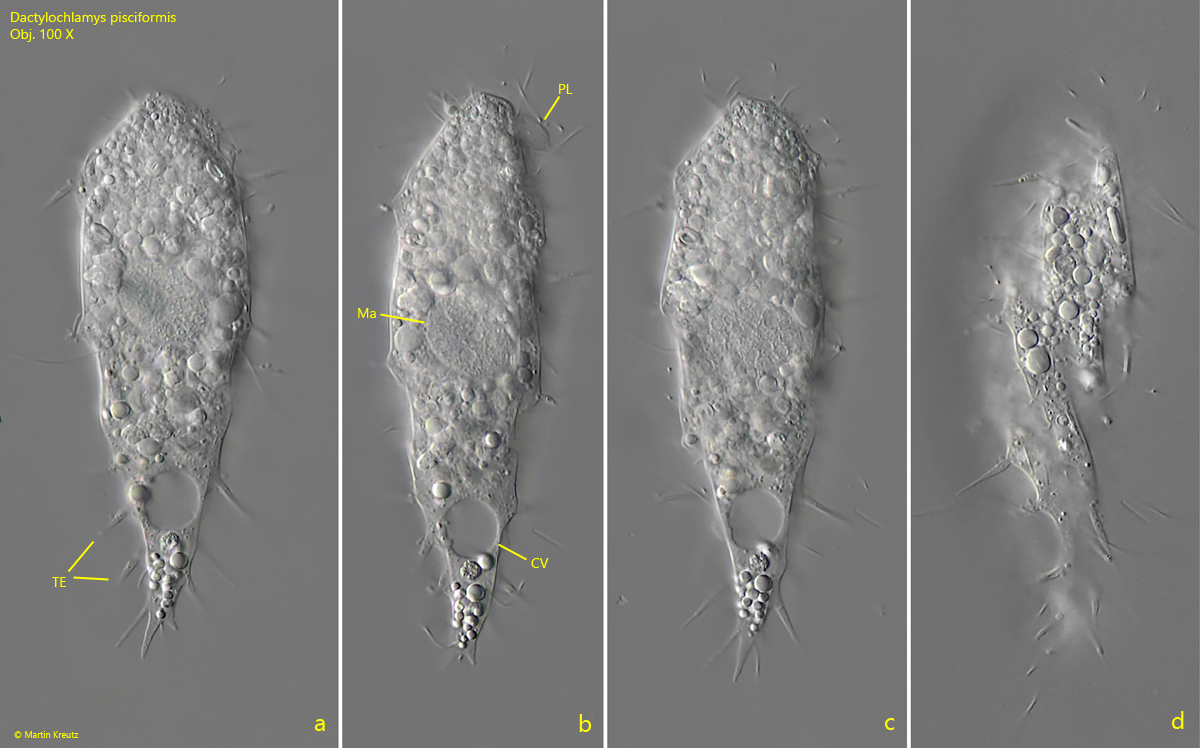
Fig. 2 a-d: Dactylochlamys pisciformis. L = 93 µm. A second freely swimming specimen. Note the distinct cytoplasm lip (PL) at the dorsal side of the oral bulge. CV = contracile vacuole, Ma = macronucleus, TE = tentacles. Obj. 100 X.
Fig. 3 a-c: Dactylochlamys pisciformis. L = 84 µm. A third, freely swimming specimen. Obj. 100 X.
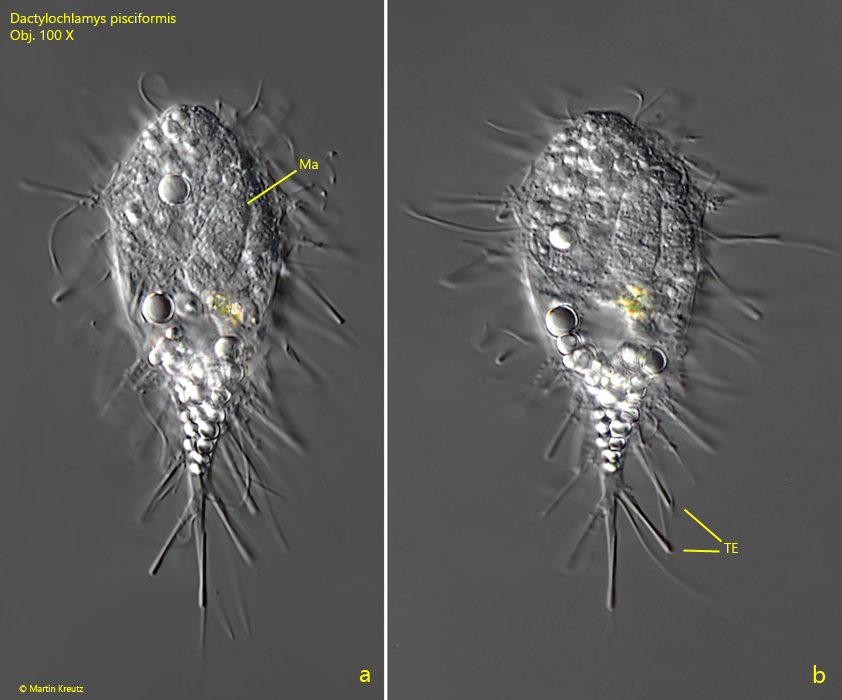
Fig. 4 a-b: Dactylochlamys pisciformis. L = 68 µm. A fourth, club-shaped specimen with partly extended tentacles (TE). Ma = macronucleus. Obj. 100 X.
Fig. 5 a-d: Dactylochlamys pisciformis. L = 68 µm. The slightly squashed specimen as shown in fig. 4 a-b. The tentacles (TE) have a slightly thickened, cylindric end. Ma = macronucleus, Mi = micronucleus. Obj. 100 X.
Fig. 6: Dactylochlamys pisciformis. L = 68 µm. A crop of fig. 5 a with details of the tentacles (TE). In each tentacle a 10 µm long, rod-shaped extrusome (EX) is located and the distal ends of the tentacles are cylindrically thickened. Obj. 100 X.
Fig. 7 a-c: Dactylochlamys pisciformis. L = 80 µm. Three focal planes of a freely swimming specimen found in October 2010 in the Suploch (Island Hiddensee). Ma = macronucleus, EX = extrusomes. Obj. 100 X.
Fig. 8 a-b: Dactylochlamys pisciformis. L = 80 µm. The slightly squashed specimen as shown in fig. 6 a-c. EX = extrusomes, Ma = macronucleus, Mi ) micronucleus. Obj. 100 X.
Fig. 9: Dactylochlamys pisciformis. In a strongly squashed specimen the 10 µm rod-shaped extrusomes (EX) are visible. Ma = macronucleus. Obj. 100 X.
Fig. 10 a-b: Dactylochlamys pisciformis. The short dorsal brush (DB) is hard to observe. The broad cilia of the dorsal brush are about 1 µm long. Obj. 100 X.
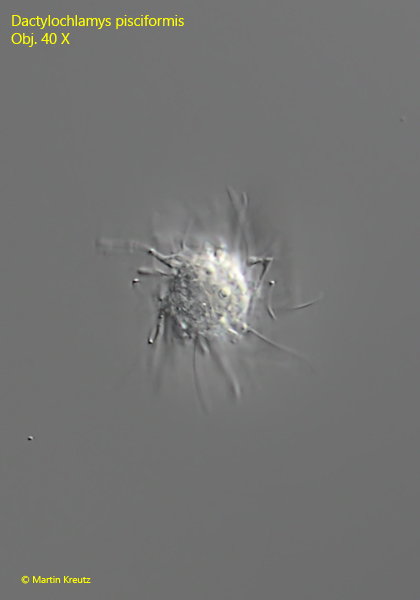
Fig. 11: Dactylochlamys pisciformis. Apical view of a freely swimming specimen. Obj. 40 X.