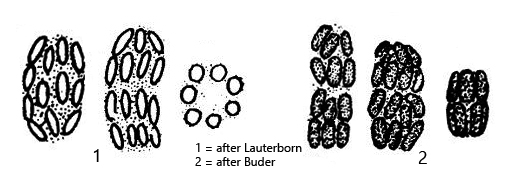
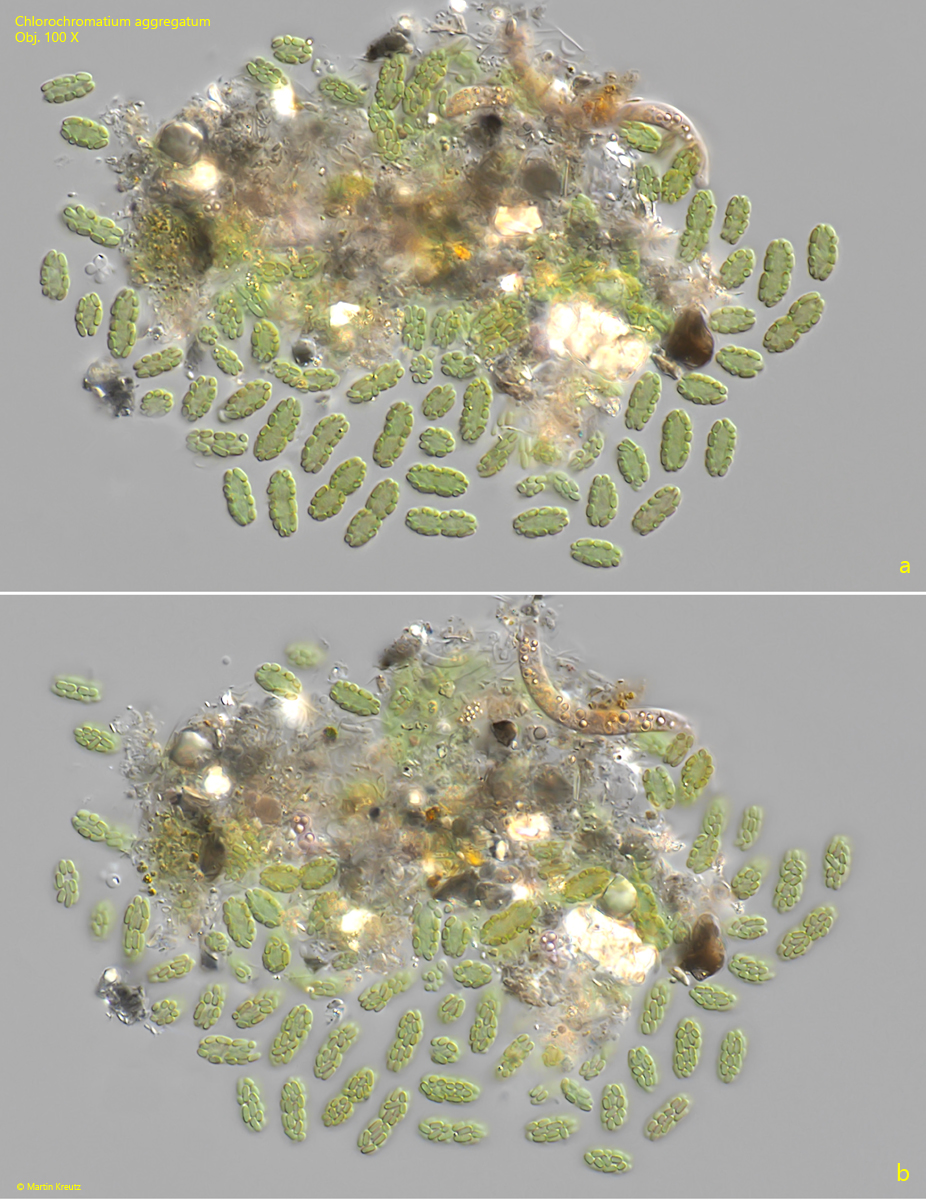
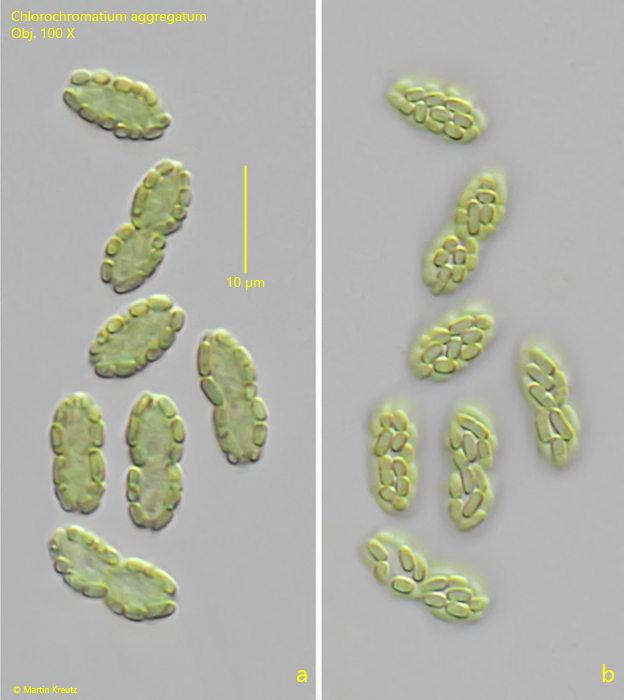
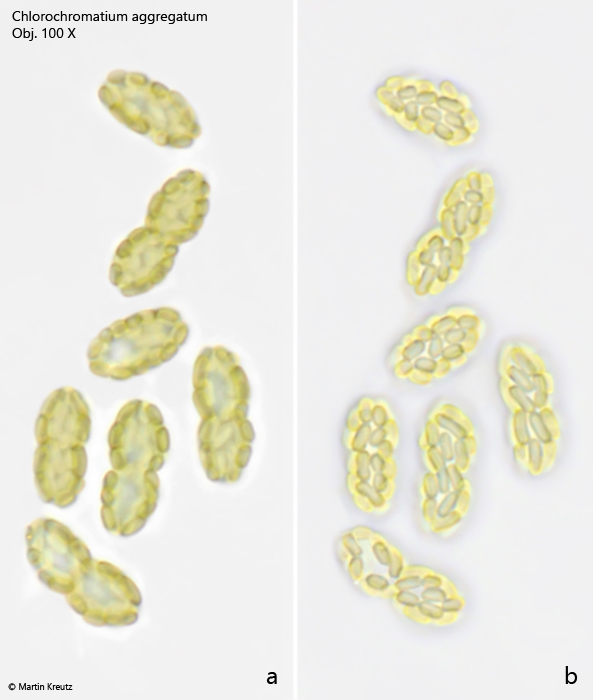
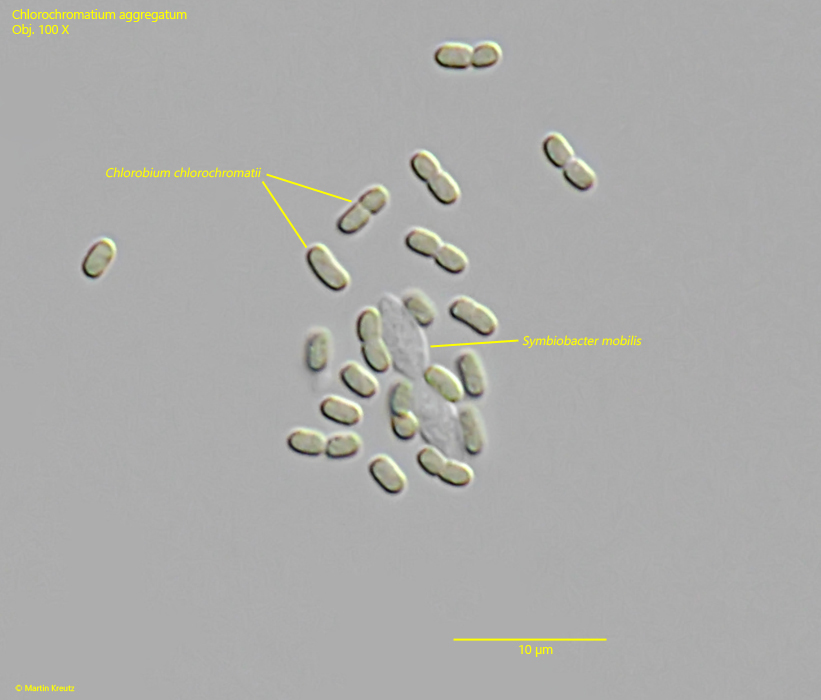
The epibiotic green sulfur bacterium was identified as Chlorobium chlorochromatium. This bacterium is capable of photosynthesis. The green-yellow coloration is caused by bacteriochlorophylls a and c (Imhoff, 1995). In culture experiments, it was demonstrated that Chlorobium chlorochromatium can be isolated and can also grow in culture without the central bacterium. Chlorobium chlorochromatium is strictly anaerobic. It uses sulfides as electron donors to fix CO2 and N2.
The central bacterium was identified by Liu et al. (2013) as Symbiobacter mobilis (referred to by the authors as “Candidatus Symbiobacter mobilis”). It possesses a polar flagellum, which makes the entire consortium motile. Symbiobacter mobilis cannot be isolated and cultured. The genome of this bacterium is so reduced that it can only survive with the help of its epibionts. It uses the photosynthetically produced products of the epibionts for energy generation. For this, Symbiobacter mobilis requires an electron acceptor. Liu et al. (2013) postulate that quinones are delivered to Symbiobacter mobilis by the epibionts, which are reduced there to hydroquinones. These are then returned to the epibionts. Thus, it is an electron cycle system.