both ends of aggregates tapered and with twisted flagella
cells hyaline, without granules or gas vacuoles
Pelosigma cohnii
I find Pelosigma cohnii very frequently in the siltation zone of the Simmelried. Near of the surface, anaerobic layers with a strong population of rhodobacteria have formed there. The aggregates of Pelosigma cohnii are usually found between these colonies of rhodobacteria.
Pelosigma cohnii was first described by Warming in 1875. Lauterborn (1913, 1915) described the species in more detail. Both authors classified Pelosigma cohnii as an aggregate of bacterial cells. About 40 years later, Skuja (1956) also found Pelosigma cohnii in the lakes of Sweden. However, Skuja interpreted and drew the aggregates as flagellates containing symbiotic bacteria (s. drawings above). It is unclear whether Skuja was aware of the work of the two earlier authors. However, it is definitely an aggregate of bacteria, as the organelles of a flagellate are also missing (e.g. nucleus or contractile vacuole). Finally Hirsch described the species again in 1974.
My population of Pelosigma cohnii differs in some aspects from those described by Skuja and Hirsch. The aggregates of my population consist of 20–50 single cells. Hirsch (1974) describes the number of single cells as 4–12.
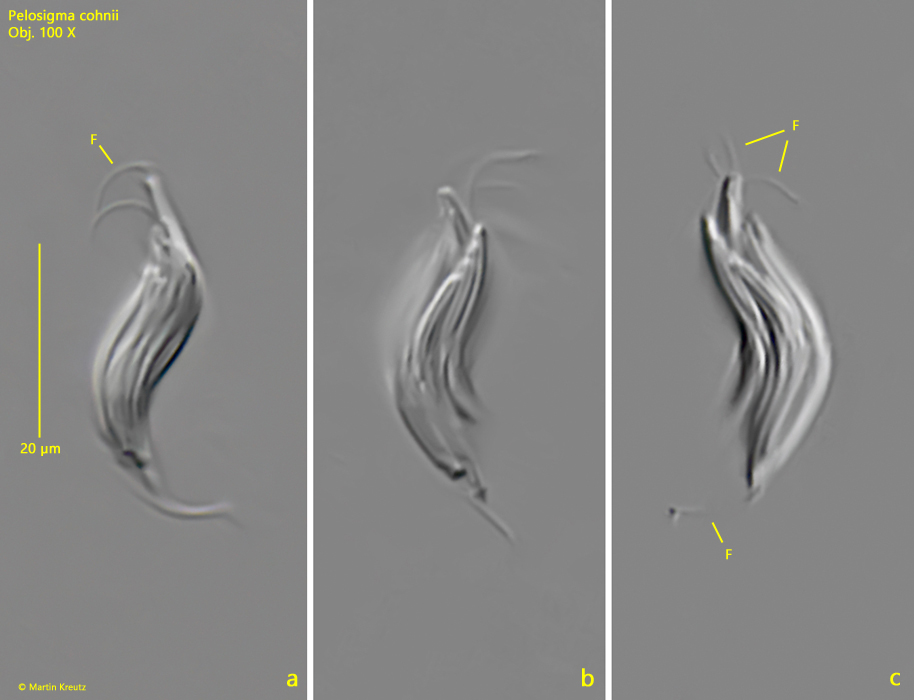
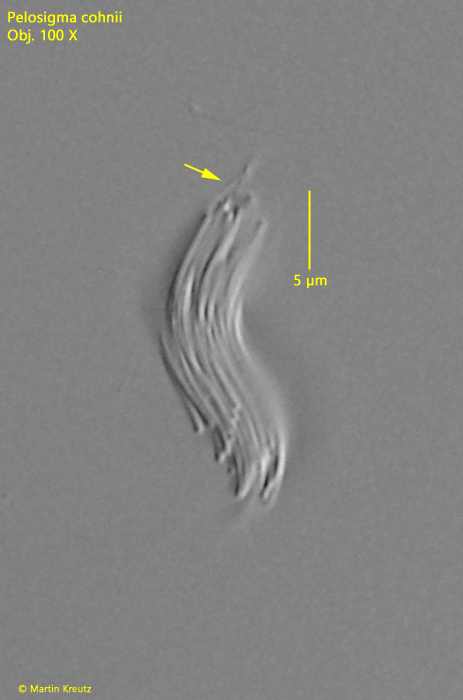
The aggregates are 15-40 µm long and tapered at both ends. Flagella arise at both ends, as described by Hirsch. The aggregates can swim slowly. The flagella appear very thick for bacteria. However, they are the twisted flagella of all the individual cells to form a compound (similar to a rope). This can occasionally be seen at maximum magnification (s. fig. 3).
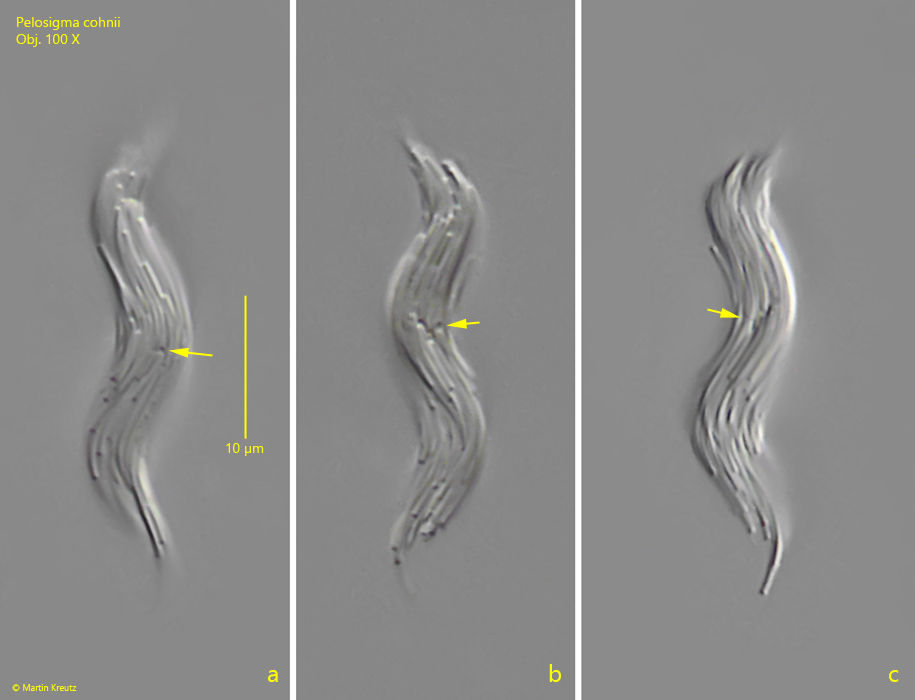
Skuja has described that the cell division occurs by longitudinal division. However, I could clearly see that the bacteria cells divide transversely as expected (s. fig. 5 a-c). This process seems to take place synchronously in the aggregates when the aggregates divide completely. This means that all the cells involved must have divided in order to divide the aggregate.
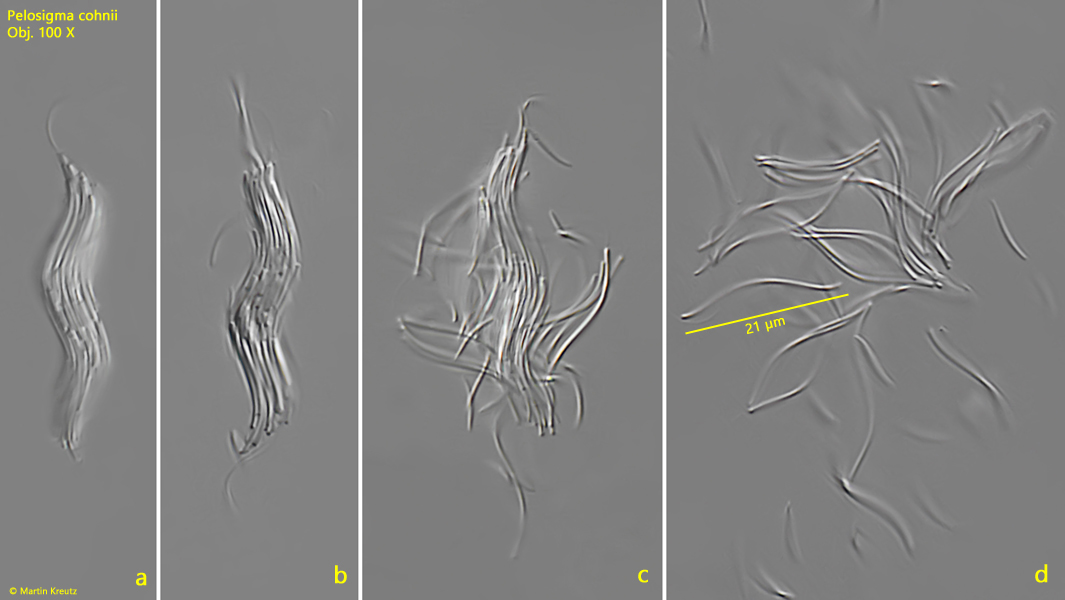
Under increased coverslip pressure, it is often possible to observe how the aggregates disintegrate into single cells (s. fig. 6 a-d). Then the sigmoid shape of the single bacteria cells can be recognized. They are about 1 µm thick and 15–22 µm long. Granules or vacuoles cannot be observed in the single cells. They appear homogeneous.
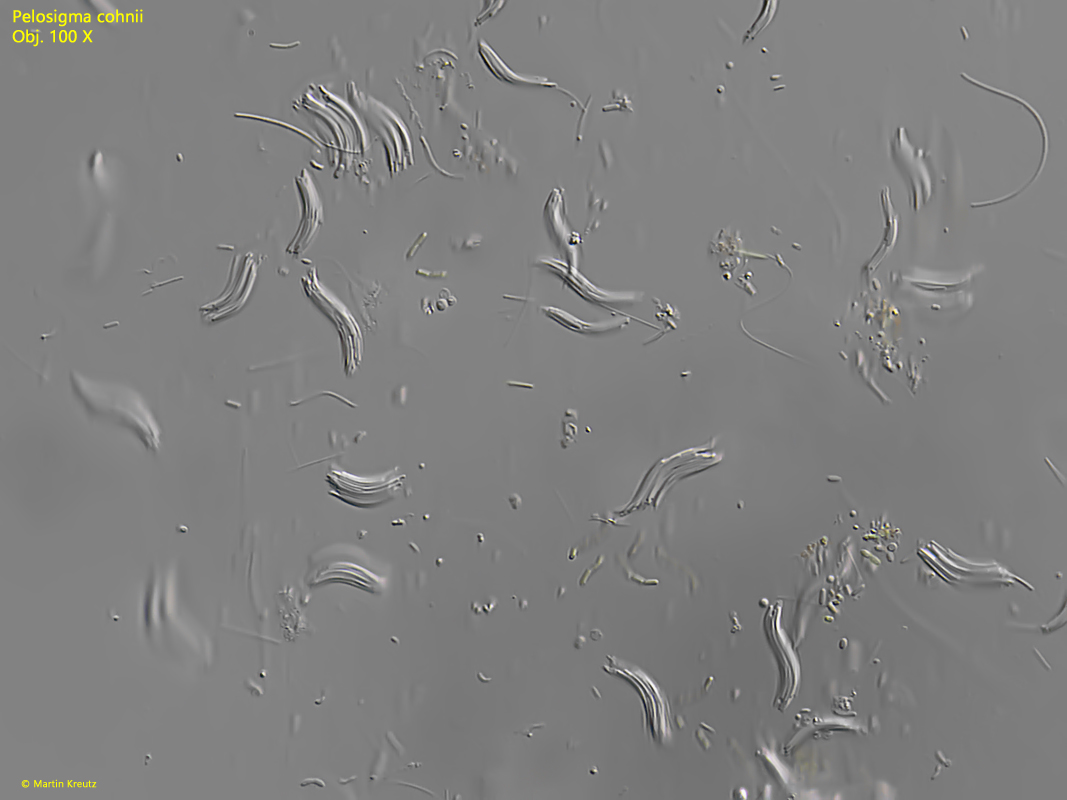
Fig. 1:Pelosigma cohnii. An accumulation of numerous sigmoid aggregates. Obj. 100 X.
Fig. 2 a-c:Pelosigma cohnii. L = 25 µm. Different focal planes of an aggregate of about 30 cells with flagella (F) at both tapered ends. Obj. 100 X.
Fig. 3:Pelosigma cohnii. L = 19 µm. The flagella of all the individual cells twist (arrow) to form a compound, which appears as a thicker flagellum. Obj. 100 X.
Fig. 4:Pelosigma cohnii. L = 19 µm. An optical cross-section through an aggregate reveals the regular arrangement of the individual cells. Obj. 100 X.
Fig. 5 a-c:Pelosigma cohnii. L = 34 µm. An aggregate during cell division. Note the division zones of the transversely dividing cells (arrows). Obj. 100 X.
Fig. 6 a-d:Pelosigma cohnii. L = 35 µm. Under increasing coverslip pressure, the aggregates begin to disintegrate into the sigmoid single cells. Obj. 100 X.